


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Как известно, внутриклеточные газовые вакуоли синезеленых водорослей способствуют вертикальным перемещениям этой группы водорослей. В условиях устойчивой температурной стратификации и недостатке фотосинтетически активной радиации поднимающиеся в освещенный эпилимнион клетки синезеленых водорослей начинают активно утилизировать диффундирующий из атмосферы в поверхностные слои углекислый газ [56, 57, 58, 59, 60]. Опускание синезеленых водорослей в нижележащие богатые биогенами слои происходит вследствие разрушения вакуолей, которые лопаются под действием увеличения давления при концентрировании в клетке углеводов при фотосинтезе. И все же синезеленые водоросли редко покидают перемешиваемый слой даже при устойчивой стратификации как по плотности, так и по биогенным веществам [61]. Причина такой «привязанности» синезеленых к перемешиваемому слою связана с зависимости миграций клеток от либо от освещенности либо от динамических явлений, типа плотностных или ветровых течений.
Соотношение толщины освещенного и перемешанного слоя воды – главная причина влияния плотностной стратификации на интенсивность процессов в стратифицированных водоемах. Толщина эпилимниона определяется взаимодействием сил плавучести (устойчивости) и динамического воздействия ветра на водную поверхность, в то время как развитие фитопланктона наблюдается только в освещенном фотическом слое, глубина которого зависит от интенсивности радиации и характеристик водной толщи (коэффициента экстинкции).
Как правило, величины этих слоев различаются, и фитопланктон может выноситься из освещенного слоя. Оптимальное развитие разных водорослей характеризуется различными соотношениями перемешанной и освещенной зон. Например, для диатомовых это соотношение значительно меньше, чем для зеленых водорослей.
Влияние стратификации на продуктивность водохранилища детально исследовано специальными наблюдениями на Можайском водохранилище [62]. Для характеристики стратификации использовалась величина квадрата частоты Вяйсяля-Брендта. В результате этих исследований установлено, что внутрисезонные колебания первичной продуктивности водохранилища зависит от сочетания температуры, устойчивости (частота Вяйсяля-Брендта) и соотношения перемешанного и фотического слоев. В последнее время лимнологи уделяют особое внимание явлению ночной конвекции, т. к по распространенному мнению именно конвекция постоянно подпитывает верхние слои минеральными биогенами даже при наличии мощного термоклина. Именно этим процессом объясняется недавно описанный «малый круговорот фосфора», обусловленный хемогенным кальцитом [65].По-видимому, именно этот механизм лежит в основе постоянного обмена биогенами внутри эпилимниона [63, 64].
В работе [66] путем моделирования описаны два пути развития «цветения» в стратифицированном водохранилище в зависимости от толщины эпилимниона. В первом случае «цветение», как указывается в большинстве работ, развивается при установлении стратификации и при относительно небольшой толщине эпилимниона. Однако, при слабой турбулентности интенсивность развития фитопланктона не зависит от толщины эпилимниона, поскольку скорость роста водорослей в освещенном слое очень высока и их биомасса слабо реагирует на вертикальное перемешивание. При этом развитие «цветение» может развиваться независимо от степени перемешивания при относительно малых глубинах (менее 15 м).
1.2.5 Гидрохимические факторы. Внешняя химическая нагрузка
Из химических факторов, влияющих на распределение фитопланктона, на первом месте, безусловно, находится концентрация химических веществ в водоеме. При этом общая минерализация может быть важным фактором видового состава фитопланктона, а концентрация питательных (биогенных) для клетки веществ, прежде всего солей азота и фосфора, продуктивности фитоценоза. Запас биогенов в водоемах обычно недостаточен для поддержания высокой интенсивности фотосинтеза, поэтому высокая продуктивность может поддерживаться только притоком биогенов из различных источников при условии их попадания в слой эпилимниона.
Суммарное поступление биогенных веществ в водоем в специальной литературе по эвтрофированию природных вод названо биогенной нагрузкой водоема. Различают площадную биогенную нагрузку равную годовой нагрузки, отнесенной к площади водоема, и объемную – отнесенную к объему водоема.
Необходимо подчеркнуть одну важную особенность водоемов замедленного водообмена ‑ скорость изменения результирующей годового внешнего баланса химических веществ в водоеме, т. е. их запаса в водных массах, значительно меньше скорости основных внутриводоемных биологических и биохимических процессов, определяющих продуктивность экосистемы. Эта особенность позволяет в проблеме оценки продуктивности водоема выделить три самостоятельные ключевые задачи, которые могут решаться независимо:
- расчет и прогнозирование поступления биогенных веществ (биогенной нагрузки) водоема;
- расчет и прогнозирование содержания биогенных веществ в водоеме с учетом процессов водообмена;
- оценка продуктивности экосистемы при данной концентрации биогенных веществ.
Вторая задача решается путем расчета по уравнениям баланса биогенов в полностью перемешиваемом водоеме. Впервые ее решил выдающийся швейцарский лимнолог Фолленвейдер, который считал, что скорость седиментации фосфора в водохранилище зависит от его общего содержания в водоеме. Фолленвейдера справедливо считают автором первого количественного описания процесса эвтрофирования озер, основанного на решении балансовых уравнений с учетом предложенной им эмпирической зависимости величины удержания фосфора от коэффициента водообмена водоема [33]. Схема системы связей в балансовых моделей Фолленвейдера представлена на рисунке 6.
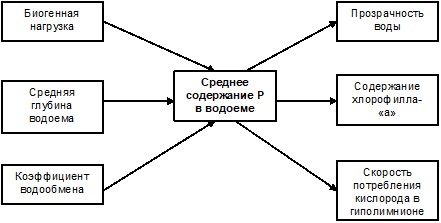
Рисунок 6 – Система связей в простых стационарных моделях эвтрофирования водоема
Биогенная нагрузка водоемов формируется из различных источников, но, главным образом, за счет приноса питательных веществ речной водой с водосбора речной системы. Поэтому наблюдается четкая зависимость первичной продуктивности водоемов от плодородия почв, типа почвы и некоторых других факторов, действующих в пределах водосбора. Водоемы, расположенные в пределах высокоплодородных почв, отличаются высоким уровнем продуктивности фитопланктона. Распашка почв и применение минеральных удобрений изменяют биологический круговорот элементов, присущей биоклиматической зоне, вызывая увеличение миграции элементов по профилю почв с нисходящим током влаги [67].
Это указывает на необходимость детального изучения процесса формирования запаса фосфора в водохранилищах в весенний период с целью прогностических расчетов развития фитопланктона в предстоящий летний период.
К числу природных абиотических факторов, определяющих биогенную нагрузку водоемов, следует отнести и атмосферные осадки [68]. В последнее время этому вопросу уделяется особенно много внимания в связи с антропогенным загрязнением природной среды. Об этом свидетельствует высокое содержание фосфора в атмосферных осадках крупных городов [69].
Химический состав и формирование запаса минеральных биогенных веществ в водной массе водохранилища может сильно варьировать от года к году в зависимости от соотношения объемов образующих ее исходных водных масс:
- основной зимней, образовавшейся осенью и трансформировавшейся в течение 4–5 месяцев под ледяным покровом благодаря процессам самоочищения;
- речной зимней;
- донной зимней, наиболее обескислороженной и насыщенной продуктами регенерации минеральных веществ из разложившегося детрита, в том числе и минеральным фосфором;
- речной весенней.
Суммарный объем первых трех масс определяется глубиной предвесенней сработки полезного объема водохранилища – чем она больше, тем этот объем меньше. А соотношение в нем объемов составляющих масс зависит от глубины слоя отбора и расхода воды при этой сработке.
Важнейшее значение для уровня развития фитопланктона в вегетационный период играет запас фосфора в водной массе водохранилища, сформировавшийся в результате его весеннего наполнения. Для установления подобной связи можно воспользоваться и концентрациями минерального фосфора, поскольку в момент наполнения водохранилища биологические процессы еще недостаточно активны и значения концентраций ортофосфатов дают определенное представление об общем запасе фосфора в водохранилище в предвегетационный период.
Сочетанием сложных внутриводоемных гидрологических процессов объясняется межгодовая изменчивость общей (внешней и внутренней) фосфорной нагрузки приплотинных и центральных районов водохранилищ в период весенней вспышки цветения преимущественно диатомовыми водорослями.
Во время летнего развития преимущественно синезеленых водорослей колебания фосфорной нагрузки в верхних районах сильно зависит от частоты паводков и их объемов, еще более изменчивых и менее предсказуемых, чем половодья. В центральных и приплотинных районах водохранилищ фосфорная нагрузка формируется весной, а затем изменяется внутримассовыми процессами обмена между трофогенным и трофолитическим слоями этой водной массы.
1.2.6 Гидрохимические факторы. Внутренняя химическая нагрузка
Если внешняя биогенная нагрузка указывает на поступление биогенов извне, то внутренняя биогенная нагрузка – на интенсивность круговорота биогенов внутри водоема. Внутренний баланс веществ отражает перераспределение отдельных химических элементов в процессе геохимической и биогеохимической миграции. Основной источник внутренней нагрузки экосистемы водоема биогенными веществами – донные отложения водоема, аккумулирующие как аллохтонный, так и автохтонный взвешенный материал. Высокая эффективность использования донных отложений в качестве индикатора экологического состояния водоема основана том, что по соотношению потоков на границе вода-дно можно относительно надежно судить о происходящих в экосистеме изменениях.
Современные теоретические представления о роли донных отложений в процессах обмена на границе вода–дно и оценки фактора биогенной нагрузки на экосистему наиболее полно обобщены в работах [70, 71] и сводятся к следующим положениям.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 |
