


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
1.2 Типы абиотических факторов развития фитопланктона в водохранилищах
1.2.1 Гидрологические факторы. Внешний водообмен
От соотношения составляющих внешнего водообмена, т. е. от структуры водного баланса, зависит интенсивность и направленность процессов круговорота вещества и энергии в водоеме, что, в свою очередь, оказывает существенное влияние на уровень продукционных процессов в его экосистеме.
Внешний водообмен определяет обмен воды водоема со смежными участками гидрографической сети, а также с атмосферой и грунтами и, соответственно, имеет горизонтальную и вертикальную составляющие. Принципиальные различия между этими составляющими имеют важнейшее значение для характера массообмена в водоеме. При увеличении вертикальной составляющей возрастает роль внутриводоемных процессов, как факторов трансформации химического стока и, следовательно, возрастает их влияние на процесс круговорота веществ в экосистеме. При доминировании горизонтальной составляющей более значима роль водосбора в формировании абиотического фона экосистемы. Соотношение вертикальной и горизонтальной составляющей зависит от структуры водного баланса водоема, которая в свою очередь определяется его географическим положением, местом в гидрографической сети, размерами и морфометрическими особенностями водоема. Один из важнейших количественных показателей, определяющих структуру водного баланса, – величина удельного водосбора. Чем она больше, тем значительней роль горизонтальной составляющей.
Нередко высказывается мнение, что замедление стока путем создания водохранилищ стимулирует основное следствие антропогенного эвтрофирования водоемов – цветение водоемов [13, 14, 15, 16]. На этом основаны и предложения по ограничению процесса эвтрофирования путем дополнительного обводнения водохранилищ, т. е. увеличения интенсивности водообмена. Как известно, при замедлении водообмена тип экосистемы изменяется с лотического (речного) на лентический (озерный) и потамофильные водные организмы постепенно вытесняются лимнофильными [17]. Теоретически, в биостатической системе увеличение биомассы водорослей лимитируется адвекцией не содержащей фитопланктон воды только в случае, если скорость удвоения биомассы оказывается меньше скорости водообмена в системе [18]. Механизм прямого влияния проточности водоема на его продуктивность состоит тогда в том, что при накоплении биомассы фитопланктона возрастает общая величина первичной продукции. Это возрастание не компенсируется естественным снижением скорости продукционных процессов, которое на самом деле имеет место и подтверждено рядом исследований [18, 19]. Удаление биомассы водорослей в результате адвекции при быстром водообмене происходит быстрее, чем образование новой биомассы, лимитированной скоростью потребления доступных биогенных элементов клетками водорослей, поэтому накопления биомассы не происходит и цветение не развивается. Таким образом, в высокопроточных водоемах биогенное лимитирование процесса первичной продукции представляется вторичным фактором по сравнению с влиянием адвекции на развитие фитопланктона.
По существующим оценкам удельной скорости роста фитопланктона [20] влияние замедления водообмена на увеличение продуктивности проявляется только в водоемах с очень высокими значениями коэффициента водообмена и уже при значении коэффициента ниже 52 год–1 (что соответствует периоду водообмена 7 суток) первичная продукция водоема становится независимой от горизонтальной адвекции. Эти теоретические выводы были подтверждены детальными наблюдениями, проведенными Д. Собалле и С. Тренкелдом в притоке и сбросе небольшого высокопроточного (средний период водообмена 5 суток) водохранилища во Флориде [18]. При очень низкой удельной скорости роста фитопланктона в исследованном ими водохранилище и высокой его проточности увеличение концентрации хлорофилла и общего содержания органического вещества в сбросе по сравнению с притоком коррелировало с проточностью водохранилища только в диапазоне периодов, при которых отношение периода водообмена к удельной скорости роста фитопланктона было больше единицы.
Следует отметить, что влияние водообмена на развитие фитопланктона заметно проявляется только в случае, когда поступающие в водоем воды характеризуются отсутствием фитопланктона или меньшим его содержанием, чем в самом водоеме. В интенсивно «цветущей» реке эффект замедления стока не увеличит степени развития планктона, если остальные факторы первичного продуцирования остаются неизменными. Таким образом, теоретически даже при очень низкой удельной скорости роста фитопланктона, порядка 0,1–0,2 сут–1 период водообмена должен быть всего одну неделю, чтобы удаление фитопланктона в результате адвекции оказывало существенное влияние на продукционные процессы. Подавляющее число озер и водохранилищ имеют период водообмена намного больше указанного значения, поэтому прямое влияние «вымывания» фитопланктона в них не проявляется.
Тем не менее, попытки получить прямые зависимости величины продуктивности водоемов от периода их водообмена, предпринятые в ряде эмпирических исследований [21, 22, 23, 24, 25, 26], привели к пороговому значению периода водообмена 60 дней, что значительно выше расчетного. Например, М. Страшкраба использовал многолетние наблюдения за продуктивностью фитопланктона в водохранилище Слапи в вегетационный период при различных значениях периода водообмена в этот период, изменяющихся в разные годы в пределах 20–100 дней, для построения зависимости первичной продукции и величины биомассы фитопланктона от периода водообмена. Зависимость, полученная М. Страшкрабой, характеризуется резко выраженным максимумом при значениях водообмена в диапазоне 40–60 дней [27]. Подчеркнем, что эти пределы влияния водообмена получены не расчетами скоростей процессов, а путем статистической обработки данных. Однако корректное сопоставление продуктивности и водообмена водоема может производиться только при полном соблюдении идентичных условий развития фитопланктона в водохранилище при различных периодах водообмена. Как уже указывалось, при увеличении периода водообмена и ослаблении горизонтальной его составляющей в водоеме заметно усиливаются процессы, связанные с внутриводоемной динамикой вод. Таким образом, изменения интенсивности внешнего водообмена неизбежно вызывают изменения ряда абиотических факторов развития фитопланктона. Эти особенности в маловодные годы при увеличении периода водообмена могут определять увеличение первичной продукции в водохранилище. Стоит добавить неизбежную несравнимость погоды при такой статистике.
Однако колебания водности года и интенсивности водообмена водоема могут приводить и к противоположному результату. Например, увеличение интенсивности водообмена в многоводные годы сопровождается увеличением притока воды в водоем, с повышенной концентрацией биогенных веществ [28, 29]. Следствием возрастания биогенной нагрузки может быть увеличение первичной продукции фитопланктона в вегетационный период. Например, Р. Тернер с коллегами [30] при изучении влияния водообмена на баланс биогенов и продуктивность небольшого водохранилища Талкин (Флорида, США) получил, что в многоводный год при высокой проточности водохранилища его продукция была в 2 раза выше, чем в маловодный. Этот результат Р. Тернер объясняет более высоким содержанием доступного фосфора в водохранилище в год с высокой проточностью, в то время как в годы пониженного водообмена более благоприятны условия осаждения сорбированного минерального фосфора, ниже его концентрация, а, следовательно, и продукция. Кроме того, при изменении интенсивности водообмена скорость образования новых клеток фитопланктона, характеризующаяся в гидробиологии временем удвоения клеток, становится более важным фактором, чем доступность биогенов. При обилии биогенных элементов в водоемах с коротким временем пребывания воды начинают доминировать быстрорастущие виды водорослей, а общая величина продукции при этом сохраняется неизменной [31].
Для подтверждения мнения о прямом влиянии внешнего водообмена на трофическое состояние экосистем иногда привлекаются диаграммы Р. Фолленвейдера. Действительно, в качестве одного из факторов, определяющих трофический уровень водоема, Р. Фолленвейдер использует величину гидравлической нагрузки (отношение годового притока воды к площади водоема), функционально связанной с коэффициентом водообмена. Но, как подчеркивал сам Фолленвейдер, гидравлическая нагрузка применена в диаграммах вместо глубины водоема для того, чтобы учесть фактор влияния внутренней нагрузки на общее поступление биогенных веществ в водоем, которая более высока в малопроточных водоемах [32]. Возрастание внутренней нагрузки в условиях замедления водообмена безусловно способствует увеличению первичной продукции в вегетационный период. Однако в этом случае речь идет лишь о косвенном, опосредованном влиянии водообмена на характер функционирования экосистем и на трофическое состояние озер и водохранилищ.
Таким образом, даже в одном водоеме связанные с изменением водности притоков межгодовые колебания интенсивности водообмена неоднозначно влияют на изменение условий для развития фитопланктона. Закономерности этих изменений изучены еще явно недостаточно и полученные в рассмотренных работах статистические связи между периодом или коэффициентом водообмена и величиной первичной продукции отражают лишь косвенное, опосредованное влияние изменения абиотических факторов на продуктивность экосистем в различные по водности годы. Если же рассматривать среднемноголетние характеристики водообмена и продуктивности различных водоемов, то связь их продуктивности с интенсивностью водообмена не выявляется. Попытка получить такую зависимость с использованием данных по характеристикам первичной продуктивности в водохранилищах России и сопредельных государств [33, 34, 35], показывает явное отсутствие связей трофических характеристик экосистемы с величиной водообмена (пример зависимости приведен на рисунке 5).
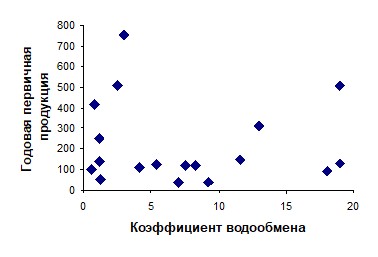
Рисунок 5 – Соотношение годовой первичной продукции (г С/м2 год) в вегетационный период (мкг/л) с коэффициентом водообмена водохранилищ (год–1) [11]

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 |
