

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Величиной, противоположной рождаемости, является смертность. Ее можно определить как число особей ΔNm, погибших за время Δt. Так же, как и при оценке рождаемости, смертность обычно относят к общему числу особей в популяции N, а промежуток времени стараются брать по возможности короче. Мгновенная удельная смертность d (death-rate, не путать со знаком дифференциала!) выражается формулой ![]() . Размерность мгновенной удельной смертности такая же, что и у рождаемости - «единица времени-1». По определению смертность может быть величиной положительной или равной 0 (последнее бывает редко и только в течение непродолжительного времени). В экологии величина смертности объединяет всех погибших особей независимо от того, умерли ли они от старости и болезней, были съедены хищником или погибли от каких-либо других неблагоприятных воздействий (отравлены пестицидами, например).
. Размерность мгновенной удельной смертности такая же, что и у рождаемости - «единица времени-1». По определению смертность может быть величиной положительной или равной 0 (последнее бывает редко и только в течение непродолжительного времени). В экологии величина смертности объединяет всех погибших особей независимо от того, умерли ли они от старости и болезней, были съедены хищником или погибли от каких-либо других неблагоприятных воздействий (отравлены пестицидами, например).
Используя введенные нами обозначения рождаемости и смертности, основное уравнение динамики численности можно записать так: r = b - d. Очевидно, что если рождаемость равна смертности, то численность остается постоянной, или, как говорят, популяция находится в стационарном состоянии. Как правило, на небольших интервалах времени b ≠ d. Понятно, что скорость изменения численности популяции по определению может быть величиной положительной, отрицательной или равной 0.
Скорость изменения численности популяции можно измерить непосредственно как изменение численности ΔN за промежуток времени Δt. Переходя к мгновенной удельной оценке, можно записать, что 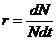 . Параметры r и b на практике оценить относительно просто. Сложнее бывает непосредственно оценить смертность d. Однако ее можно определить окольным путем по разности между рождаемостью и скоростью изменения численности.
. Параметры r и b на практике оценить относительно просто. Сложнее бывает непосредственно оценить смертность d. Однако ее можно определить окольным путем по разности между рождаемостью и скоростью изменения численности.
6.3. Продолжительность жизни, демографические таблицы и кривые выживания
Для каждого организма характерна некая максимальная продолжительность жизни. Эта величина варьирует очень широко: от десятков минут у бактерий (время между делениями) до нескольких тысячелетий у некоторых деревьев (секвойя Sequoiadendron giganteum и сосна Pinus longaeva, С. Америка), т. е. в диапазоне 103 - 1011 с.
Обычно продолжительность жизни крупных животных и растений больше чем мелких (почти линейное возрастание), но и здесь встречаются исключения (рукокрылые живут до 30 лет).
Вполне понятно, что продолжительность жизни организмов одного вида может сильно меняться в зависимости от условий существования. Например, пойкилотермные животные, как правило, живут дольше при низкой температуре, чем при высокой (если эти температуры не выходят за пределы толерантности).
Как ни странно, но разговор о росте численности популяции проще начинать с анализа смертности, в частности с рассмотрения того, как смертность в тех или иных популяциях распределяется по возрастам. Такую информацию, т. е. сведения о распределении смертности по возрастам, обычно представляют в виде таблиц выживания.
В ряде случаев самый простой способ построения таких таблиц – это подробное наблюдение за судьбой когорты – небольшой группы особей, отрожденных в популяции за короткий промежуток времени, и регистрация возраста наступления смерти всех членов данной когорты. Рассмотрим, как строятся подобные когортные таблицы на примере данных Дж. Коннела (Connel, 1970), который наблюдал в течение 10 лет за когортой усоногого рачка Balanus glandula в приливно-отливной зоне одного из островов у северо-западного побережья США. Свои наблюдения Коннел начал в 1959 г., спустя приблизительно 1-2 месяца после того, как произошло оседание личинок, а закончил в 1968 г. году, когда погибли последние особи из выбранной когорты (таблица).
· В первой графе указан возраст х (в данном случае в годах);
· Во второй – число особей, доживших до этого возраста: nx;
· В третьей – доля особей, доживших до определенного возраста: lx = nx/no, где no – число особей начального возраста;
· В четвертой: число особей dx, погибших в течение интервала от x до x + 1;
· В пятой – смертность qx в течение интервала х: qx = dx/nx.
В таблицу могут входить и другие параметры, например, т. н. ожидаемая продолжительность жизни и т. п.
Рассмотренная таблица называется когортной, поскольку построена по данным наблюдений за динамикой смертности в одной конкретной когорте. Составить такие таблицы можно только в случаях, когда имеется реальная возможность проследить за индивидуальной судьбой всех членов достаточно большой выборки из определенной когорты. Сравнительно легко это было сделать для популяции балянусов, каждый из которых пожизненно прикреплен к скале, но и в этом случае понадобились регулярные наблюдения на протяжении 10 лет. Собрать аналогичный материал для организмов, ведущих подвижный образ жизни, или для организмов, характеризующихся большой продолжительностью жизни, гораздо труднее. Однако существует другой способ построения таблиц выживания.
Вместо того чтобы наблюдать за отдельной когортой в течение периода, близкого по времени к максимальной продолжительности жизни, исследователь может в течение небольшого периода времени наблюдать за смертностью в отдельных возрастных группах (т. е. в сосуществующих когортах), а, зная численность этих групп, рассчитать специфическую для каждого возраста смертность. Таким способом нередко пользуются демографы, поскольку проследить на протяжении почти столетия за судьбой по крайней мере тысячи людей достаточно сложно, да, пожалуй, и невозможно.
Таблица выживания, построенная на основании краткосрочных наблюдений за смертностью во всех возрастных группах, называется статической. Статическая таблица представляет собой как бы временной срез через популяцию.
Если мы построим график зависимости доли доживших особей lx от возраста х, то получим кривую, называемую кривой выживания (дожития), или просто «lx-кривой». Выделяют три основных типа кривых выживания (рисунок):
1-й тип (сильно выпуклая кривая) – отражает ситуацию, при которой смертность до определенного возраста ничтожно мала, а затем быстро возрастает и все особи погибают за короткий срок. Такой тип характерен для имаго многих насекомых, для некоторых ящериц, африканских копытных, для большинства млекопитающих, в том числе и человека;
2-й тип (диагональная кривая) – свидетельствует об относительном постоянстве смертности всех возрастов. Другими словами это означает, что в течение каждого возрастного интервала гибнет одна и та же доля от численности когорты в начале этого интервала. Этот тип свойствен преимущественно птицам, но встречается и у многих других животных, например, у гидры, некоторых ящериц, у бородавочника;
3-й тип (сильно вогнутая) – очень высокая смертность в раннем возрасте, сменяющаяся относительно низкой смертностью оставшихся особей. Данный тип кривой выживания иногда называют типом устрицы (Ostrea), поскольку у этого моллюска, ведущего во взрослом состоянии прикрепленный образ жизни, есть планктонная личинка и именно на стадии личинки наблюдается чрезвычайно высокая смертность особей. Такой тип кривой характерен практически для всех организмов с большой плодовитостью и отсутствием заботы о потомстве. К числу таковых относятся практически все морские и пресноводные беспозвоночные, рыбы, а также большинство насекомых, если рассматривать не имаго, а весь онтогенез.
Реально встречающиеся кривые выживания нередко представляют собой некоторую комбинацию указанных выше «основных» типов. Например, характерная для крупных млекопитающих кривая I типа почти всегда в самом начале круто падает вниз, что соответствует значительной смертности сразу после рождения.
Чтобы получить полную картину динамики численности той или иной популяции, а также рассчитать скорость ее роста, наряду с данными о том, как распределяется по возрастам смертность, необходимо знать также, в каком возрасте особи начинают производить потомство и какова средняя плодовитость каждого возраста. Для этого можно построить новую таблицу. Для простоты представим себе гипотетический пример животного, начинающего размножаться на третьем году жизни и живущего, как правило, не более 10 лет. В первой графе таблицы запишем возраст (х), во второй – долю особей, доживших до данного возраста от начальной численности когорты (lx), в третьей – среднее число потомков, появившихся на свет у особей данного возраста в расчете на одну родительскую особь (mx), в четвертой – произведение доли доживших особей на среднюю их плодовитость (lx mx). Сумма всех значений последнего столбца называется чистой скоростью воспроизводства:
![]()
Чистая скорость воспроизводства показывает, во сколько раз увеличивается численность популяции за одно поколение. Если R0 = 1, то популяция стационарна – численность ее сохраняется постоянной, поскольку каждое последующее поколение точно замещает предыдущее. Очевидно, что величина R0 значительно варьирует в зависимости от вида организмов, а также от условий его существования.
7. ЭКСПОНЕНЦИАЛЬНЫЙ РОСТ ПОПУЛЯЦИЙ
7.1. Построение модели экспоненциального роста
Рассмотрение модели экспоненциального роста популяций начнем с примера с мускусным овцебыком. За период с 1700 по 1800 гг. эти крупные животные в результате охоты были почти полностью истреблены в пределах всего своего естественного ареала обитания в Северной Америке. В Аляске, например, последние особи были убиты в 1850-1860 гг. В 1930 г. власти Аляски спохватились и закупили небольшое стадо из Гренландии для повторного вселения овцебыков. Животных разместили на одном из достаточно больших островов, где отсутствовали хищники, охотящиеся на овцебыков, и где для них было достаточно корма. В 1936 г. на остров высадили 31 животное. Цель этого мероприятия заключалась в том, чтобы по мере роста популяции «лишних» животных изымать и расселять по новым территориям.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
