

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
В период с 1936 по 1947 гг. проводились более или менее регулярные учеты численности популяции овцебыка. Полученные данные представлены на рисунке. Попытаемся использовать эти данные для построения математической модели, описывающей рост популяции овцебыка.
Вспомним, что общее число особей любого вида (N) на некоторой ограниченной площади может определяться только такими факторами, как рождаемость, смертность, иммиграция и эмиграция. Для простоты изложения материала здесь и в дальнейшем будем использовать не мгновенные, а дискретные изменения численности популяции:
![]()
Чтобы подчеркнуть, что в этом уравнении используются дискретные величины рождаемости, смертности, эмиграции и иммиграции, будем обозначать их заглавными латинскими буквами. В случае с овцебыком период времени, за который происходили изменения численности популяции, равен 1 году. В случае с другими видами, эти временные промежутки, конечно, будут меняться. Очевидно, что за год численность популяции изменится на величину, равную 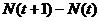 .
.
Многие популяции, подобно рассматриваемой популяции овцебыка на острове, являются закрытыми, т. е. в них отсутствуют такие процессы, как иммиграция и эмиграция. Поэтому для таких популяций приведенное выше уравнение запишется так: ![]() .
.
Мы уже говорили, что рождение и гибель особей представляют собой разнонаправленные процессы, которые характеризуются определенной интенсивностью. В целом соотношение процессов рождения и гибели особей можно выразить при помощи специфической скорости роста популяции. Опять-таки, чтобы не путать дискретную скорость роста популяции с мгновенной (r), обозначим эту величину буквой R.
По величине R можно судить не только о росте, но и о снижении численности популяции. Очевидно, что если R > 1, то численность популяции растет. Наоборот – при R < 1 численность будет снижаться. Наконец, в ситуации, когда число родившихся особей равно числу погибших (R = 1), численность популяции не будет изменяться.
Если мы захотим предсказать численность популяции овцебыка через два года, то записанное выше уравнение придется использовать дважды:
![]()
Объединив эти два уравнения, получим:
![]()
Такая запись означает, что для нахождения численности популяции через t лет необходимо исходную численность умножить на параметр R t раз.
Последнее уравнение и представляет собой экспоненциальную модель динамики численности популяции. Если использовать мгновенную скорость роста популяции r, то уравнение это можно записать так: ![]() . Иногда этот тип роста называют еще геометрическим, или мальтузианским (в честь Томаса Мальтуса).
. Иногда этот тип роста называют еще геометрическим, или мальтузианским (в честь Томаса Мальтуса).
7.2. Допущения экспоненциальной модели
При построении любой модели приходится вводить определенные допущения, которые упрощают реальную картину изучаемого явления или процесса и на 1 место ставят наиболее важные факторы, определяющие эти процессы или явления. Естественно, это касается и модели экспоненциального роста популяций. Рассмотрим допущения этой модели.
1. Модель экспоненциального роста – это полностью детерминистическая модель. Это означает, что она однозначно предсказывает, чему будет равна численность популяции через промежуток времени t. При этом не учитываются всевозможные колебания факторов среды, которые, естественно, будут вносить значительную неопределенность в предсказание численности популяции. Одним из проявлений этой неопределенности было то, что рассчитанная нами скорость роста популяции была неодинакова в разные годы, и лишь колебалась около некой средней величины.
2. Предполагается, что популяция растет по экспоненте в течение бесконечно длительного времени. Однако, как мы увидим далее, такая картина в природе может наблюдаться лишь непродолжительное время и только в условиях, когда популяции не ограничена ресурсами и в ней нет внутривидовой конкуренции за эти ресурсы.
3. Предполагается, что рождаемость и смертность особей не зависит от их возраста или каких-то других уникальных характеристик. Т. е. допускается, что все особи в популяции совершенно одинаковы. Однако очевидно, что в реальных популяциях вероятность выживания и размножения, способности к эмиграции и иммиграции у разных особей будут в значительной степени варьировать. На это будут оказывать влияние такие факторы, как возраст, пол, размер, здоровье, социальное положение, генетические особенности особей. Однако даже если рождаемость и смертность зависят от возраста, то оказывается, что в пределах определенных возрастных групп эти параметры в среднем оказываются постоянными. Поэтому если предположить, что соотношение возрастных групп в популяции постоянно, то и скорость роста самой популяции будет постоянна, о чем и говорит экспоненциальная модель.
4. Предполагается, что популяция является панмиктической, и что особи в ней распределены равномерно. Взаимодействие с другими популяциями того же вида характеризуются процессами эмиграции и иммиграции, интенсивности которых постоянны во времени. Эффекты популяций других видов на изучаемую популяцию также считаются постоянными (например, одинаковое во времени давление хищников). Очевидно, однако, что в природных популяциях все эти условия выполняются лишь в определенной небольшой степени.
8. ФАКТОРЫ, РЕГУЛИРУЮЩИЕ РОСТ ПОПУЛЯЦИИ.
ЛОГИСТИЧЕСКАЯ МОДЕЛЬ РОСТА ПОПУЛЯЦИИИ
8.1. Факторы, регулирующие рост популяции
Как мы уже отмечали, экспоненциальный рост происходит в благоприятных для популяции условиях. Однако рано или поздно среда становится не такой благоприятной; например, популяция может пережить несколько холодных зим подряд, в результате чего численность потомства снизится. Для других видов такими неблагоприятными событиями могут быть засуха (или наводнение) или любые другие факторы, которые ответственны за колебания в природе. Подобные климатические и другого рода изменения, наступление которых обычно предсказать невозможно, называются регуляторными факторами, независимыми от плотности популяции.
Тем не менее, большее значение для ограничения экспоненциального роста, по-видимому, имеет ограничение популяции по какому-то ресурсу или ресурсам, возникающее вследствие постепенного их исчерпания. Такими ресурсами могут быть вода, пища, биогенные элементы, солнечный свет, пространство и некоторые другие. Поскольку особи одной популяции пользуются одинаковыми ресурсами, то по мере роста численности популяции, на каждую особь начинают приходиться все меньшие количества этих ресурсов. Рассмотрим основные последствия этого снижения доступа к ресурсам:
1) Рост смертности. Особи многих видов при возрастании численности популяции начинают переселяться в новые, менее заселенные области. Однако некоторые виды не обладают механизмами активной эмиграции. Так, у большинства растений семена распространяются при помощи ветра или животных. В результате одним из основных последствий перенаселенности у таких видов становится возрастание уровня смертности. Существование такого эффекта было доказано экспериментально. Здесь можно привести пример с травянистым растением Cakile edentula, произрастающим на песчаных дюнах. В эксперименте применили разную плотность посадки семян этого растения – от 1 до 200 на 400 см2. Выживаемость растений оказалась в строгой обратно пропорциональной зависимости от плотности посадки (выживаемость рассчитывалась как доля растений, сформировавших зрелые семена).
2) Снижение плодовитости. Когда особи получают меньше ресурсов, интенсивность их размножения может существенно снижаться. Так, у птиц величина кладки в значительной степени определяется доступностью пищи. Показано например, что при увеличении численности популяции большой синицы Parus major, обитающей в одном из парков Оксфорда, с 40 до 90 особей, средняя плодовитость снизилась с 4,5 до 2,5 оперившихся птенцов на гнездо.
3) Истощение особей. Высокая плотность популяции обычно сопровождается также снижением средних размеров и индивидуальных весов особей. В логарифмическом масштабе подобная зависимость выглядит в виде прямой линий, угловой коэффициент которой, как правило, равен -1,5.
4) Изменения территориального поведения. Вспомним: территория представляет собой определенный участок пространства, охраняемый одной, двумя, или несколькими особями от посягательств со стороны других членов популяции. Территориальное поведение особенно характерно для птиц. При увеличении плотности популяции животных с территориальным поведением возможны следующие исходы: а) взаимное расположение территорий может стать более плотным; б) территории могут стать меньшими по размеру; в) некоторые животные могут быть изгнаны в менее подходящие местообитания; г) наконец (хотя это последний эффект), может значительно возрасти эмиграционная активность животных. Часто лишенные территории особи не питаются. В результате в популяции снижается средняя плодовитость, даже если плодовитость остальных, кормящихся особей остается на том же уровне.
Помимо рассмотренных эффектов перенаселенности, растущая популяция может отравлять среду обитания собственными отходами жизнедеятельности, а также становиться более доступной для хищников и болезней. Все перечисленные факторы, т. е. нехватка «пищи» в широком понимании, усиление воздействия «врагов» (хищники, болезни), отравление среды называются факторами, зависимыми от плотности. В совокупности эти факторы объясняют, почему экспоненциальный рост численности популяции на протяжении длительного времени невозможен.
8.2. Логистический рост популяции

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 |
