
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Хордотимпальные слуховые органы состоят из струнообразной связки, натянутой над покровами тела, и сенсорных клеток, которые одним концом соединяются со связкой, а другим – с покровами. Связка колеблется в унисон звуковым волнам, поступающим из окружающей среды, и возбуждает рецепторные клетки.
Орган слуха позвоночных развивается вместе с органом равновесия из утолщения эктодермы в задней части головы. Этот зачаток углубляется и принимает вид пузырька, который в дальнейшем разделяется на 2 отдела. Из верхнего отдела образуются полукружные каналы и овальный мешочек, а из нижнего формируется полое выпячивание – лагена и круглый мешочек. Все эти образования заполнены жидкостью – эндолимфой и составляют так называемый перепончатый лабиринт. Он окружен стенками скелета, а промежуток между ними и перепончатым лабиринтом заполнен перилимфой.
У круглоротых, рыб и безногих амфибий со слуховой рецепцией связаны 2 сосочка круглого мешочка. Для амфибий характерен амфибийный сосочек, а для остальных тетрапод – базилярный сосочек, в которых звуковоспринимающую функцию выполняют рецепторные волосковые клетки.
У рептилий, птиц и однопроходных имеется вытянутый слепой вырост – лагена. У птиц она к тому же изгибается, а у млекопитающих многократно закручивается, образуя улитку. У млекопитающих литка разделяется тремя перепонками на 3 части.
Две из них – это лестница преддверия и барабанная лестница, содержат перилимфу и соединяются между собой на верхушке улитки через небольшое отверстие. Средний канал улитки (перепончатый лабиринт) содержит эндолимфу. Он отделен от барабанной лестницы базилярной мембраной, а от лестницы преддверия – рейснеровой. Базиллярная мембрана состоит из поперечно ориентированных коллагеновых волокон («струн»), длина которых по направлению к основанию улитки постепенно уменьшается. Эти волокна резонируют в унисон звукам различной высоты.
|
Рис. 20. Схема поперечного разреза улитки, на котором виден кортиев орган. |
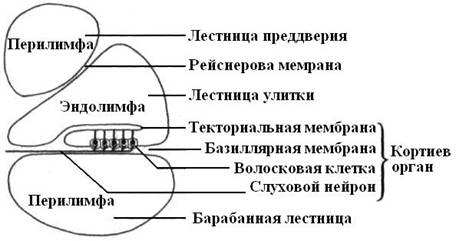
Располагающийся на базилярной мембране звуковоспринимающий кортиев орган состоит из сенсорных волосковых и поддерживающих эпителиальных клеток, которые подразделяются на наружные и внутренние.
На апикальных участках сенсорных клеток имеются пучки коротких уплотненных микроворсинок, способных отклоняться – это стереоцилии. Стереоцилии наружных сенсорных клеток своими вершинами прикасаются к поверхности лежащей над ними текториальной мембраны. Наружные сенсорные эпителиоциты значительно чувствительнее к звукам большей интенсивности, чем внутренние.
|
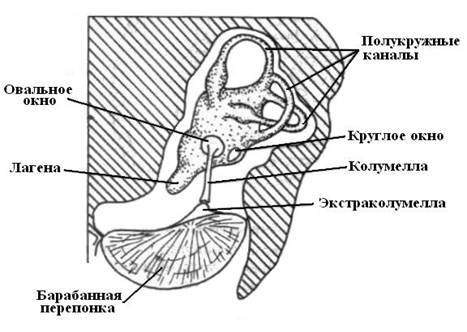
Рис. 21. Организация слуховой системы птиц. |
Первичноводные позвоночные довольно широко используют коммуникации, связанные с испусканием звуков. Очень часто они для этого используют растяжение плавательного пузыря. Звуковоспринимающую функцию у них выполняют рецепторные клетки макул круглого мешочка и лагены. Звуковые волны эффективно передаются в воде, поэтому у первичноводных животных нет специализированных образований для фильтра и усиления звуков. Эту функцию выполняет все тело, по тканям которого колебания воды передаются практически без потерь.
У наземных позвоночных условия звукопередачи на внутреннее ухо изменяются, т. к. воздействие звуковой волны оказывается слишком слабым для рецепторного аппарата внутреннего уха. Поэтому у них в эволюции сформировались специальные приспособления для усиления звуковых сигналов – среднее и наружное ухо. Все особенности структур среднего уха направлены на усиление внешних звуковых сигналов.
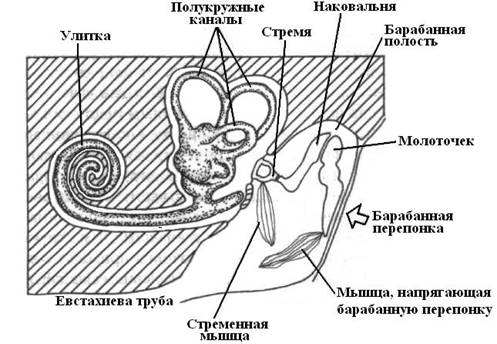
Вся система среднего уха сложилась на основе морфогенетических изменений одной из висцеральных щелей – брызгальца, наружная часть которого несколько расширилась, образовав барабанную полость, и закрылась барабанной перепонкой, а внутренняя сузилась и превратилась в евстахиеву трубу – узкий канал, конец которого, как и брызгальца открывается в глотку. В среднем ухе имеется набор косточек, которые передают колебания от барабанной перепонки к овальному окну. У амфибий, рептилий и птиц сигналы передаются через столбик (колумелла), а у млекопитающих – через наковальню, молоточек и стремечко.
|
Рис. 22. Организация слуховой системы млекопитающих. |
У змей и некоторых других рептилий барабанная перепонка редуцирована. У них колебания воздуха передаются от квадратной черепа через короткую связку колумелле, а от нее к овальному окну.
Наружное ухо присуще птицам и млекопитающим. Птицы обладают слухом в большом частотном диапазоне, что обеспечивается организацией рецепторного аппарата и особенностями наружного уха. У птиц лагена в сравнениями с рептилиями увеличена в размерах, но еще не формирует завитков, как у млекопитающих. Слуховые рецепторные клетки у птиц располагаются в лагене в перепончатом лабиринте, который во многом похож на лабиринт млекопитающих.
У млекопитающих слуховой аппарат внутреннего уха представляет собой длинный рукав, который закручен в виде улитки. Исключение составляют однопроходные (яйцекладущие), у которых практически прямой рукав внутреннего уха (нет улитки). Полость среднего уха соединяется с ротовой полостью евстахиевой трубой и отличается главным образом большой сложностью передаточного аппарата, состоящего из трех слуховых косточек. Особая мышца, прикрепляющаяся к рукоятке молоточка, натягивает барабанную перепонку, а другая – мышца стремени, наклоняет стремечко и натягивает перепонку овального окна. Рефлекторно сокращением этих мышщ уменьшается амплитуда колебаний всего аппарата при более сильных звуках.
У млекопитающих значительного развития достигает наружный слуховой проход и связанные с ним наружные ушные раковины. Последние собирают звуковые волны и направляют их в проход. Ушная раковина имеет хрящевой скелет и приводится в движение особой мускулатурой. У китообразных в связи с их образом жизни ушная раковина отсутствует, наружный слуховой проход редуцирован, и звуки проводятся к среднему и внутреннему уху через жировые и костные ткани черепа.
Одной из основных характеристик слуха является восприятия звуков определенного диапазона частот. Наибольшее разнообразие в этом отношении характерно для позвоночных животных. Так, рыбы чутко реагируют на инфразвуковые и низкочастотные колебания, издаваемые плавающими в воде животными. Сами рыбы издают с аналогичным диапазоном звуки типа: хрюканья, крика, тарахтенья, гудков, скрежета и др. Пресмыкающиеся слышат звуки более высокого тона, чем амфибии. Большинство птиц наиболее чувствительны к звукам 3,0 – 3,5 кГц, собаки – 35 кГц, летучие мыши и дельфины до 100 кГц, человек от 16 до 18 кГц. В то же время тимпанальные органы кузнечиков слышат звуки около 200 кГц.
Согласованная деятельность обоих ушей обеспечивает по разности силы и фазовому сдвигу звуковых колебаний, приходящих в одно и другое ухо, определять направление к источнику звука.
Аномалии развития органа слуха у человека.
Встречается полное отсутствие ушной раковины и наружного слухового прохода, малая или, наоборот, большая их величина. Возможно недоразвитие внутреннего уха с атрофией слухового нерва.
Фоторецепторные системы.
Для животных свет – это один из наиболее эффективных видов дистантной рецепции. Живые организмы приспособились к восприятию световых лучей, достигающих поверхности Земли в диапазоне от 200 до 1200 нм.
Фоторецепторные системы появляются уже у кишечнополостных – у медуз. У них фоторецепторные клетки развиваются из эктодермы и сосредотачиваются по краю зонта и на щупальцах. У червей светочувствительные клетки рассеяны по всей поверхности тела, но у некоторых из них на головном конце тела они образуют скопления, окруженные пигментом. Эти, так называемые «глазки», позволяют животным ориентироваться по направлению света. У моллюсков возникает оптический аппарат, превращающий поток света в изображение, что дает начало предметному зрению. Оно совершенствовалось путем эволюции фасеточного и камерного глаза. Кроме того, многие беспозвоночные сохранили рассеянные по поверхности тела светочувствительные клетки.
Реже фоторецепторные клетки лежат на одном уровне с покровным эпителием (например, у медуз). Чаще они погружены в глазную ямку, что предохраняет их от грубых механических раздражений. В ряде случаев поверх рецепторных клеток формируется прозрачная кутикула, обеспечивающая еще более надежную защиту. Если края ямки смыкаются, то образуется пузырчатый (камерный) глаз. В таких случаях прозрачные клетки передней стенки глазного пузырька превращаются в хрусталик, который отбрасывает изображение предметов окружающего мира на фоторецепторные клетки, что дает начало предметному зрению. Большую роль при этом играет пигмент, который изолирует каждую светочувствительную клетку от побочных лучей света за исключением тех, которые падают на нее в строго определенном направлении. Без такой оптической изоляции образное зрение было бы невозможным.
Дальнейшее усовершенствование пузырчатого глаза сводится к увеличению числа фоторецепторных клеток и приобретению диафрагмы – зрачка, устраняющего боковые лучи, а также к появлению аппарата аккомодации, позволяющего глазу настройку на рассмотрение дальних или близких предметов. Аккомодация достигается или перемещением хрусталика или изменением его кривизны.
Глаза камерного типа, где и оптические структуры и фоторецепторные клетки располагаются в одной камере, дают сплошное, но обратное изображение увиденного. Их имеют паукообразные и моллюски, особенно головоногие. Но наибольшего совершенства такие глаза достигают в эволюции позвоночных животных и у человека.
У членистоногих бывают глаза двух типов – простые и сложные (или фасеточные). Простой глаз состоит из единственной линзы, прикрывающей небольшое число фоторецепторных клеток. Такие глаза могут различать свет и темноту, но не способны к предметному зрению.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 |
