
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
|
Рис. 23. Схема строения отдельного омматидия. А – продольный разрез; Б – поперечный разрез. |
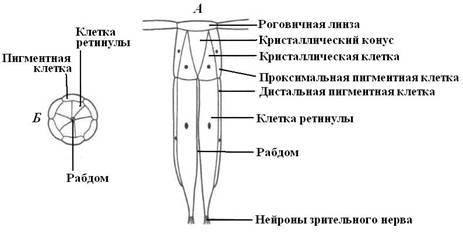
Фасеточные глаза состоят из тысяч отдельных зрительных единиц – омматидий (рис. 24).
Каждый омматидий имеет две светопреломляющие структуры – роговичную линзу и кристаллический конус, являющийся производным особых клеток. Каждый омматидий обрабатывает изображение только одной точки рассматриваемого предмета. Линза роговицы, образованной прозрачной кутикулой, посылает пучок света на кристаллический конус, где он проходит через отверстие, окруженное пигментными клетками, а затем попадает на фоторецепторные клетки, нервные импульсы от которых попадают в головной ганглий. Изображение прямое, но фасеточное. Фоторецепторы насекомых избирательно чувствительны к тому или иному цвету, но, кроме того, они чувствительны к ультрафиолетовому участку спектра. Например, глаз таракана лучше всего реагирует на зеленый и ультрафиолетовый цвет.
Важным свойством зрения насекомых является способность к прослеживанию быстрых движений, достигающих нескольких сотен в секунду. Фасеточные глаза приспособлены к видению, как на ярком свете, так и в сумерках благодаря особому механизму адаптации фоторецепторов к изменению освещенности.
Камерный глаз имеют некоторые беспозвоночные, однако наибольшего развития глаз такого типа достигает у осьминога, позвоночных и человека.
Глаз состоит из трех оболочек: 1) склеры (фиброзной оболочки), которая в передней части глаза становится прозрачной и образует главную линзу – роговицу; 2) сосудистой оболочки, которая содержит пигмент и представлена собственно сосудистой оболочкой, ресничным телом и радужкой; 3) аккомодационным аппаратом – хрусталиком, обеспечивающим фокусировку изображения на сетчатке; 4) сетчаткой, ретиной содержащей фоторецепторные и нервные клетки. Наполненное водянистой влагой пространство перед радужной оболочкой и хрусталиком называется передней камерой глаза, а позади радужной оболочки – задней. За хрусталиком полость глаза заполнена желеобразным стекловидным телом.
|
Рис. 24. Развитие глаза. А, Б, В – сагиттальные разрезы эмбрионального зачатка глаза на различных стадиях развития: 1 – эктодерма; 2 – хрусталиковая плакода – будущий хрусталик; 3 – глазной пузырек; 4 – сосудистая выемка; 5 – наружная стенка глазного бокала – будущая сетчатка; 7 – стебелек – будущий зрительный нерв; 8 – хрусталиковый пузырек. |
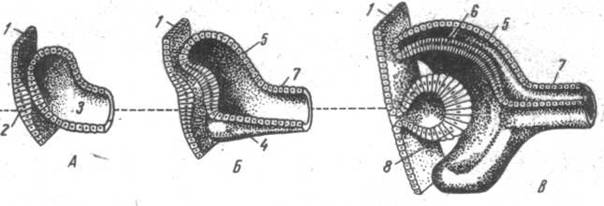
Эта общая схема строения глаза сохраняется практически во всех группах позвоночных, что отражено в его эмбриональном развитии. Зачаток глаза появляется у эмбриона как пара неглубоких желобков в выростах переднего мозга. Позже эти выросты образуют глазные пузыри, которые связаны с эмбриональным мозгом при помощи глазных стебельков. Глазные пузыри вступают в контакт с эктодермой будущей лицевой части головы и индуцируют в ней развитие хрусталика. Инвагинация стенки глазного пузыря приводит к формированию двухслойного глазного бокала, из наружного слоя которого развивается пигментный слой сетчатки, а из внутреннего – сетчатка. Сосудистая оболочка формируется из окружающей глаз белой мезенхимы. Склера, цилиарная мышца образуются тоже из мезенхимы. Зачаток хрусталика отделяется от эктодермы. Из последней развивается и эпителий роговицы.
Общий план строения глаза не исключает многообразия организации. У многих крокодилов и сумчатых млекопитающих в пигментной оболочке накапливается гуанин, образующий зеркальную прослойку – тапетум. Аналогичная отражающая поверхность имеется у рыб и млекопитающих в сосудистой оболочке. Ее появление связано с необходимостью повышения освещенности фоторецепторного слоя. Свет, который проходит через сетчатку, отражается от зеркальной поверхности тапетума и возвращается к фоторецепторам. Для многих акул и костистых рыб характерно изменение отражающей способности зеркального слоя в зависимости от освещенности.
В глазах рептилий и птиц имеются специфические образования сосудистого происхождения – папиллярный конус и гребень. У рептилий папиллярный конус не связан с фоторецепцией, а участвует в метаболизме внутренней части глаза. Для птиц характерно разнообразное строение гребня, на котором располагается сетчатка.
У позвоночных сетчатка имеет инвертированный характер: ее фоторецепторы расположены на задней дальней от зрачка поверхности сетчатки и развернуты светочувствительными сегментами в сторону пигменного эпителия. Свет проходит через всю толщу сетчатки и тела самих фоторецепторов, прежде чем попадет на зрительный пигмент мембраны дисков дистальных отростков фоторецепторных клеток.
У многих млекопитающих глаза расположены фронтально, чем достигается бинокулярное зрение. В таких случаях зрительные поля обоих глаз перекрываются, что имеет ряд преимуществ, по сравнению с использованием каждого глаза в отдельности. Чем больше глаза направлены вперед, тем шире поле стереоскопического зрения. У человека, например, общее поле зрения охватывает 180°, а стереоскопическое – 140°. У животных, у которых глаза расположены по бокам головы, фронтальное стереоскопическое поле бывает ограниченным и используется лишь для рассмотрения удаленных предметов. Для того, чтобы лучше рассмотреть близкий предмет, животное поворачивает голову и использует монокулярное зрение.
Стереоскопическое зрение позволяет получить более точное представление о размерах и форме предметов, а также о расстоянии, на котором они находятся. В основном стереоскопическое зрение характерно для хищных животных. У животных, которым приходится спасаться от хищников, глаза, наоборот располагаются по бокам головы, благодаря чему они имеют более широкий обзор, но ограниченное стереоскопическое зрение, так, например, у кролика общее поле зрение составляет 360°, а стереоскопическое – всего 20°.
Аномалии развития глаза у человека.
Глаз развивается из многих зачатков, поэтому встречаются различные эмбриональные нарушения развития глаза.
Частым дефектом является формирование неправильной кривизны роговицы и хрусталика – астигматизм, в результате чего изображение проецируется в виде не точки, а линии. Из-за неправильного формирования глазного яблока может быть врожденная близорукость (длинный глаз) или дальнозоркость (короткий глаз), не корригирующиеся преломляющими средами глазного яблока и требующие ношения соответствующих очков.
При врожденном помутнении хрусталика возникает препятствие прохождению светового пучка. В результате недоразвития венозной пазухи, склеры и пространства радужно-роговичного угла развивается глаукома, обусловленная повышением внутриглазного давления, что приводит к атрофии светочувствительных и нервных элементов сетчатки.
Пороком развития вспомогательных органов глаза является отсутствие век, а при скрытом глазе глазная щель будет весьма узкой. Встречается нависающая горизонтальная складка верхнего века. Очень редко наблюдается порок развития, когда в сосудистой оболочке отсутствует пигмент (альбиносы).
Электрорецепция и электрические органы.
У первичноводных позвоночных часть специализированных механорецепторов боковой линии трансформировалась в электрорецепторы. Помимо первичноводных позвоночных электрорецепция обнаружена у утконоса, и есть предположения о наличии такого рода рецепторов у птиц. Известно около 70 видов рыб с развитыми электрорецепторами, кроме того, около 500 видов могут генерировать электрические разряды. А, примерно, 20 видов обладают способностями и генерировать и рецептировать электрические поля.
По способности генерировать и рецептировать электрические сигналы животных делят на три группы. К первой группе относят рыб, генерирующих сильные заряды (электрические угри, электрические скаты и африканские электрические сомы). Ко второй группе относят животных, которые способны как генерировать, так и рецептировать электрические сигналы (гимнархи и клювокрылые). К третьей группе относят животных, способных только чувствовать электрические разряды, но не способных их генерировать. К этой группе относят кошачьих и молотоголовых акул. Чувствительность их электрорецепторов очень высокая, и это позволяет им определять место укрытия рыб в песке по локальному искажению электромагнитного поля Земли.
Органы, генерирующие электрические разряды имеют различное происхождение. Одни из них являются видоизмененными мышцами, а другие - нервными окончаниями. При этом электрические органы, сформированные из мышц, обычно являются низкочастотными или разряжающимися одиночными разрядами. В тех случаях, когда электрические органы формируются из нервных окончаний, частота разрядов бывает в несколько раз выше.
При формировании электрорецепторных органов из мышц последние утрачивают сократительные элементы и принимают уплощенную форму. Как правило, они имеют асимметричные потенциалы, которые складываются в силу того, что сами электрорецепторы образуют комплексы, в которых последовательно соединяются. Для синхронизации работы этих клеток существует специальный нервный аппарат, который находится под контролем среднего, заднего и продолговатого мозга, а у электрического сома контроль за синхронизацией электрических разрядов осуществляется на уровне спинного мозга. Есть и другие варианты синхронизирующих отделов мозга.
|
Рис. 25. Ампульные и клубочковые электрорецепторы. А - кожи акул; Б – костных рыб. |
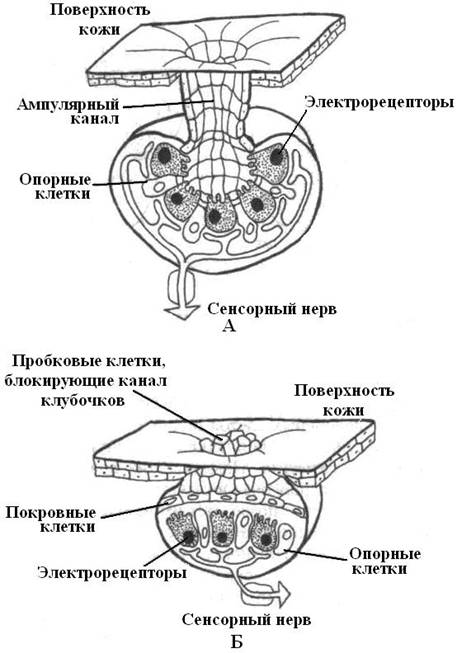
Среди электрочувствительных рецепторов выделяют два типа: ампульные и клубочковые. У некоторых видов водных позвоночных присутствуют оба типа, а у большинства – один из типов.
Ампульные электрорецепторы приспособлены для восприятия медленно изменяющихся электрических полей, а клубочковые реагируют на быстрые. Электрические сигналы, достигнув рецепторных клеток ампулы или клубочка, преобразуются в рецепторный потенциал волосковой клетки, откуда сигналы передаются к окончаниям сенсорного нерва по средством химического синапса. Следует заметить, что клубочковые рецепторы менее чувствительны, чем ампулярные.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 |
