
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
У трехслойных беспозвоночных гонады развиваются из мезодермы. При этом яйцеклетки и сперматозоиды созревают в разных железах: первые – в яичниках, вторые – в семенниках. У плоских червей часть женской гонады обособляется и специализируется на образовании не гамет, а клеток, заполненных питательным веществом, необходимым для трофики яйцеклеток. Появляется и сложная система выведения половых продуктов из гонад: яйцеводы, матка, влагалище. Наряду с яичниками формируются и многочисленные семенники, где происходит сперматогенез, и система семявыносящих каналов.
Приблизительно такая же организация половых желез и системы каналов, выводящих половые продукты, характерна и для более высокоорганизованных беспозвоночных. Однако, в отличие от позвоночных половые железы у них не выполняют гормонообразующей функции, а специализируются только на гаметогенезе.
В некоторых случаях половая система зародыша может дифференцироваться не в соответствии с генетически предопределенным полом. Так, например, в случае тестикулярной феминизации при наличии у человека мужского генотипа (46ХУ) фенотипически формируется женский пол. У таких лиц отмечается отсутствие клеточных рецепторов тестостерона. В результате этого, несмотря на нормальный уровень циркулирующего тестостерона, он не оказывает влияния на образование мужских половых органов.
Нарушение половой дифференцировки может проявляться в мужском или женском псевдогермафродитизме. Мужской псевдогерматофрадитизм наблюдается при множестве эндокринных расстройств (дефекты синтеза тестостерона, его метаболизма и эффектов на клетки – мишени). При этом при генотипе 46ХУ имеются яички, но маскуминизация неполная: может быть гипоспадия (локализация наружного отверстия полового члена или на его нижней поверхности, или в области мошонки, или в области промежности), недоразвитие полового члена, плохо развитая мошонка с яичками, или без них.
Женский псевдогермафродитизм является следствием чувствительности плода с ХХ генотипом к воздействию мужских половых гормонов во время критического периода (8-12 недели внутриутробного развития). Характеризуется увеличением клитора, наличием урогенитального синуса, выраженным губомошоночным сращением и др.
При эндокринных дисфункциях половых желез происходит нарушение полового созревания, которое сопровождается либо задержкой полового созревания либо преждевременным половым созреванием. Задержка полового созревания у девочек выражается в отсутствии вторичных половых признаков к 14-ти летнему возрасту и отсутствием менструаций к 16-ти годам при наличии вторичных половых признаков.
У мальчиков о задержке вторичных половых признаков свидетельствует отсутствие признаков полового созревания к 14-ти летнему возрасту.
Половое созревание считается преждевременным, если какой-нибудь из вторичных половых признаков появляется у девочек ранее 7,5 лет, а у мальчиков ранее 9 лет.
Аномалии развития половой системы, связанные с расстройствами гормональной функции половых желез.
Расстройства гормональной функции половых желез в эмбриональном периоде могут приводить к аномалиям развития внегонадных половых структур, а также к нарушениям половой дифферинцировки. У ранних зародышей млекопитающих, в том числе и человека, в ходе развития половой системы формируются два протока с потенциальными возможностями превращения в определенные органы мужской или женской половой системы. Это мужской (вольфов, или мезонефрастический) проток и женский (мюллеров, или парамезонефрический). Под влиянием гормона семенников – тестостерона – мюллеровы протоки развиваются в семявыносящие пути, соединяющие семенники с мочеиспускательным каналом. Кроме того, под действием тестомтерона закладываются связанные с семявыводящими протоками семенные пузырьки и предстательная железа. Потенциально женские мюллеровы протоки дегенерируют под действием другого гормона семенников зародышей – мюллерова ингибирующего фактора. В отсутствие тестостерона вольфоровы протоки дегенерируют, а отсутствие мюллерова ингибирующего фактора приводит к развитию парамезонефротических протоков в яйцеводы, матку и часть влагалища.
Наружные половые органы также закладываются в морфологически индифферентном виде, но в дальнейшем развиваются в мужские (под действием тестостерона) или в женские (если отсутствует влияние тестостерона) гениталии.
Последняя стадия половой дифференцировки начинается после рождения – в пубертатном (подростковом) периоде, когда появляются вторичные половые признаки характерные для мужского или женского пола. Это развитие определяется специфическим воздействием половых гормонов: для женского пола – гормоны яичников, для мужского – гормоны семенников.
Эндокринный аппарат поджелудочной железы хордовых.
Поджелудочная железа - это орган смешанной секреции, поскольку содержит как экзокринную часть, секретирующую панкреотическй сок, так и эндокринный аппарат.
У ланцетника и круглоротых поджелудочная железа еще не имеет органного типа, но у них присутствуют железистые клетки, гомологичные экзо - и эндокриноцитам поджелудочной железы более высокоорганизованных животных. У ланцетника такого типа клетки располагаются между эпителиоцитами слизистой оболочки кишки и печеночного выроста (в дивертикуле), образуя эндоэпителиальную железу мозаично-клеточного типа (рис. 42). Эндокринные клетки неоднородны: одни из них синтезируют и выделяют гормоны глюкагон и гастрин, а другие – инсулин.
Вышеуказанные типы эндокринных клеток одним концом достигают просвета кишечника, а другим вступают в контакт с базальной мембраной, под которой располагаются кровеносные капилляры.
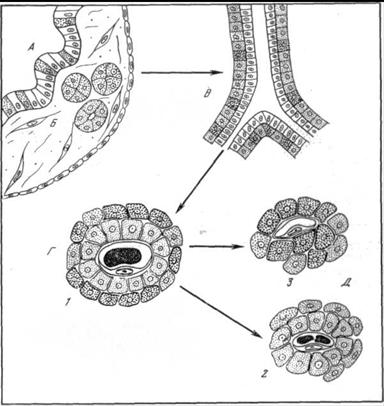
Рис.42. Схема этапов эволюции форм структурной организации эндокринной части поджелудочной железы позвоночных.
А – инсулярный аппарат бесчерепных (железа мозаично-клеточного типа); Б – инсулярный аппарат круглоротых (скопления и фолликулы Лангерганса); В – эндокринная часть поджелудочной железы хрящевых и кистеперых рыб («внешний эпителий»); Г – эндокринная часть поджелудочной железы костистых рыб, земноводных, пресмыкающихся, птиц и млекопитающих («смешанные» панкреатические островки); Д – эндокринная часть поджелудочной железы птиц («темные» и «светлые» панкреатические островки); 1 – А – клетка; 2 – В – клетка; 3 – Д –клетка
( по Яглову, 1978).
У круглоротых железистые клетки, гомологичные инсулярному аппарату поджелудочной железы более высокоорганизованных позвоночных, располагаются как в соединительнотканном слое самой кишки, так и в соединительной ткани, окружающей кишечник. Причем, в некоторых случаях описаны скопления клеток инсулярнного аппарата даже в печеночной паренхиме. Прямая связь эндокриноцитов с капиллярным руслом кишечника полностью завершается только у взрослых животных. Выделяют два основных скопления эндокринных клеток – краниальное (в начале передней кишки) и каудальнаое (в области впадения жёлчного протока).
Эмбриональным источником развития инсулярного аппарата у круглоротых является кишечный эпителий клетки, которого уже на ранних стадиях развития начинают мигрировать в собственный соединительнотканный слой слизистой оболочки. И хотя на данной стадии развития еще отсутствует прямая связь эндокринных клеток инсулярного аппарата с капиллярным руслом кишечника, еще в личиночном периоде начинается его функционирование. Окончательное формирование эндокринного аппарата завершается у взрослых особей после метаморфоза.
Одновременно с миграцией эндокринных клеток из кишечного эпителия идет процесс васкуляризации инсулярного аппарата, который полностью завершается только у взрослых особей. В центральных участках скоплений эндокринных клеток появляются заполненные коллоидоподобным веществом полости, и таким образом формируются фолликулы, получившие названия фолликулов Лангерганса.
Клеточный состав инсулярного аппарата неоднороден. В нем имеются гранулярные и агранулярные клетки. Основная масса гранулярных клеток представлена В-клетками, образующими инсулин. Роль других гранулярных клеток не ясна. Агранулярные клетки, видимо, осуществляют усвоение гормонов из коллоида фолликулов Лангерганса и обеспечивают их транспорт в капиллярное русло. Однако, это предположение остается недоказанным. Нет данных и о наличии А – клеток, образующих глюкагон.
Впервые поджелудочная железа приобретает органный статус у хрящевых рыб. У них это компактный орган дольчатого строения, состоящий из экзокринного и эндокринного отделов. Эндокринные клетки располагаются на ходу средних и мелких выводных протоков. У некоторых видов хрящевых рыб имеются другие формы организации эндокринной части железы: у одних эндокринные клетки окружают выводные протоки их в виде эпителиального слоя («внешний эпителий»); у других, кроме этого, имеются скопления эндокринных клеток в виде прилежащих к выводным протокам столбов; у третьих – помимо эндокринных клеток, располагающихся вокруг протоков, встречаются изолированные островки таких клеток.
У хрящевых рыб в отличие от круглоротых впервые появляются три вида самостоятельных гормонопродуцирующих клеток - А, В, и Д. У подкласса цельноголовых описан еще и четвертый тип клеток, названный Х – клетками. Гормональный профиль идентифицирован только для А - , В - и Д - клеток. Клетки А продуцируют глюкагон, В-инсулин, а Д-соматостатин. Последний оказывает ингибирующий эффект на экзокринный отдел железы и на выделение гормонов А - и В-клетками.
У костных рыб поджелудочная железа имеет характер компактного органа только у хищных, а у остальных рыб она представлена гроздьями железистой ткани, разбросанными в брыжейке и других органах.
У латимерии имеются скопления эндокринных клеток как по ходу средних и мелких выводных протоков, а также в виде свободно расположенных островков, не связанных с выводными протоками. Эндокринный отдел представлен А - , В - и Д-клетками.
У двоякодышащих рыб эндокринная часть поджелудочной железы имеет вид только островков А - и В - эндокриноцитов. Такой же тип организации эндокринного отдела характерен и для костистых рыб. Клеточный состав эндокринных островков у них изучен недостаточно: в них идентифицировано от одного до шести видов секреторных клеток. У большинства изученных видов идентифицированы А -, В - и Д - клетки.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 |
