
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Особенности морфо-функциональных преобразований
головного мозга человека после рождения.
К моменту рождения головной мозг, хотя в общих чертах похож на мозг взрослого человека, остается еще не полностью сформированным. Его масса составляет всего 370-392 г., к шести месяцам она удваивается, к трем годам - утраивается, а к девятилетнему возрасту в среднем достигает 1300 г., то есть всего на 100 г. меньше массы головного мозга взрослого человека.
У новорожденного основные извилины больших полушарий уже сформированы, однако в дальнейшем в соответствии с увеличением массы и размеров головного мозга происходит утолщение коры, изменяется форма, глубина и высота борозд и извилин. Окончательную форму они приобретают между 7 – 14-ю годами. Развитие цитоархитектоники головного мозга в основном осуществляется до 13 лет, хотя структурная перестройка коры в определенной степени происходит на протяжении всей жизни.
Развитие проводящих путей интенсивно продолжается до 14 – 15 лет.
Важным этапом в развитии мозга является миелинизация нервных волокон. Она начинается на 4 – 5-ом месяцах эмбрионального развития и продолжается после рождения, заканчиваясь в переднем, промежуточном и среднем мозге к двум годам, в мозжечке – к четырем, а в продолговатом мозге – к семи годам.
Промежуточный мозг к моменту рождения хорошо развит, ядра гипоталамуса вполне дифференцированы, эпифиз и гипофиз тоже хорошо развиты. К шестому месяцу жизни ребенка нервные волокна таламуса покрываются миелином.
Средний мозг принимает строение подобное тому, что характерно для взрослого организма, к 5 – 6-ти годам, однако, до 7 лет его водопровод остается более широким, а пигментация в красных ядрах и черной субстанции заканчивается к 16-ти годам.
Он имеет неглубокие борозды и меньшую массу. После рождения начинается его рост, а с 3-х месяцев – дифференцировка клеточных структур. Его относительные азмеры приближаются к мозжечку взрослого в промежутке от 1 до 2 лет. Миелинизация нервных волокон белого вещества мозжечка заканчивается к 4-м годам.
|
Рис. 12. Развитие головного мозга человека. 1 – головной мозг 2-месячного эмбриона. 2 – головной мозг 5-месячного эмбриона. 3 – головной мозг новорожденного. 4 – головной мозг взрослого человека. |
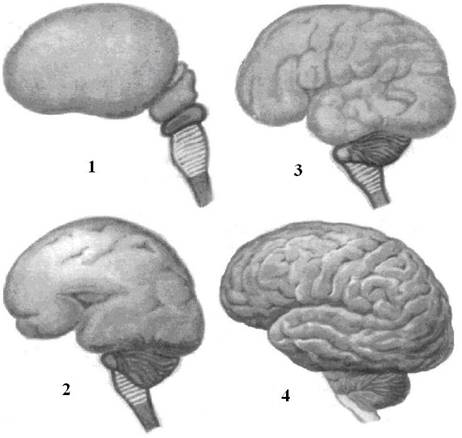
Продолговатый мозг в функциональном отношении к моменту рождения является в значительной мере созревшим, однако, миелинизация нервных волокон в нем заканчивается только к 7-ми годам.
В соответствии с морфологическим созреванием головного мозга происходит и развитие высшей нервной деятельности ребенка. К моменту рождения мозг ребенка уже готов к образованию временных связей. Есть основания полагать, что рефлекторные дуги становятся достаточно сформированными еще до рождения уже на основе пищевых безусловных рефлексов. На 15-й день после рождения удается выработать условный рефлекс на положение тела в ответ на сложный комплекс кожных, проприоцептивных и лабиринтных раздражителей.
Образование первых условных рефлексов протекает относительно медленно, а сами они еще неустойчивы, что обусловлено, видимо, широкой иррадиацией в коре процессов возбуждения и торможения. И только с 5 месяцев оказываются налицо все основные физиологические механизмы, лежащие в основе высшей нервной деятельности.
Если в первые дни после рождения проявляются первые безусловные ориентировочные рефлексы, то начиная с 3 – 4-х месяцев происходит образование условных ориентировочных (исследовательских) рефлексов, играющих в дальнейшем важную роль в поведении ребенка.
С конца первого года жизни ребенка и весь второй год происходит становление второй сигнальной системы – речи, а вместе с этим условные рефлексы начинают формироваться не только на базе безусловного подкрепления, но и на основе речи. Однако, в этом возрасте рефлексы еще не устойчивы. Посторонние раздражители у значительной части детей тормозят условные рефлексы. Это говорит о слабости процессов возбуждения и торможения в коре головного мозга.
Начиная с трехлетнего возраста вторая сигнальная система все более приобретает доминирующее значение. У детей 4-5 лет условно-рефлекторные связи при речевом и оборонительном подкреплении образуются легче, чем на пищевом подкреплении.
С 6-летнего возраста дети начинают пользоваться понятиями, абстрагированными от действия. В этом возрасте приступают к обучению письму и чтению.
К 7 годам жизни происходит морфологическое созревание лобного отдела головного мозга. В этот период появляется способность удерживать программу действия из нескольких движений и предвидеть результаты действия.
Для детей 7-9-летнего возраста характерно быстрое упрочение положительных и отрицательных условных рефлексов, их устойчивость к внешним воздействиям, более быстрая концентрация нервных процессов.
Начиная с 10-12 лет условные рефлексы на простые и сложные раздражители появляются быстро и сразу становятся прочными. В то же время условно-рефлекторные связи могут быть легко переделаны.
В подростковом возрасте (у девочек в 12-15 лет, а у мальчиков в 13-16 лет) явно преобладают процессы возбуждения, причем не только в коре, но и в подкорке. Поэтому ответные реакции могут носить неадекватно бурный характер. Речь становится более медленной, ответы носят лаконичный характер. Условные рефлексы на непосредственные сигналы образуются быстрее, а на словесные – медленнее. Корковые процессы характеризуются нарастанием возбуждения и ослаблением всех видов внутреннего торможения.
Начиная приблизительно с 15 лет роль второй сигнальной системы вновь возрастает. Возбудительные процессы в коре и в подкорке начинают снижаться, усиливается внутреннее торможение. Подростки вступают в юношеский возраст с относительно гармоничным отношением двух сигнальных систем, коры и подкорки.
Филогенез вегетативной (автономной)
нервной системы у хордовых.
Первые признаки вегетативной нервной системы намечаются у ланцетника, у которого из дорсальных корешков нервной трубки отходят нервы, идущие к диффузным сплетениям нейронов во внутренних органах. В последних еще нет специализированных ганглиев. Есть предположение, что это признаки парасимпатической системы.
У круглоротых имеются коллатеральные ганглии, которые собирают часть висцеральных волокон, расходящихся от ганглиев к внутренним органам. Однако симпатическая цепочка ганглиев еще отсутствует.
У позвоночных значительного развития достигает симпатический отдел нервной системы. Они состоит в основном из нервных узлов расположенных около позвоночного столба и связанных продольными тяжами нервных пучков.
Вегетативная нервная система хорошо развита у хрящевых рыб. Для нее характерно отсутствие коллатеральных ганглиев, расчлененность симпатической цепочки и несвязанность с ней черепных нервов. В ряде черепных и туловищных нервов симпатической системы переключение происходит в самом иннервируемом органе, как в парасимпатической системе. Большая часть симпатических нервов имеет короткие преганглионарные волокна, которые оканчиваются в симпатических узлах, лежащих вдоль спинного мозга, и длинные постганглионарные волокна, достигающие иннервируемых органов.
У костистых рыб симпатический нервный ствол непрерывен и связан с черепными нервами. Для большинства костистых рыб характерно появление коллатерального ганглия. В их голову начинают проникать туловищные волокна, которые иннервируют ротоглоточную полость и желудок. Из туловищного отдела спинного мозга выходят также нервы, иннервирующие внутренние органы. При этом они имеют короткие преганглионарные и длинные постганглионарные волокна. В каудальной части туловища часть преганглионарных волокон длинные, а постганглионарных короткие. Последние располагаются в стенке мочевого пузыря, что похоже на парасимпатическую систему млекопитающих.
Для амфибий и других тетрапод характерна парная симпатическая цепочка ганглиев, идущая вдоль спинного мозга, и многочисленные коллатеральные ганглии, расположенные между внутренними органами.
У рептилий и птиц, кроме паравертебрального симпатического ствола, выделяют непарные ганглии: верхний и нижний брыжеечные и чревный (брюшной). Их часто называют превертебральными. Большая часть симпатических ганглиев удалена от иннервируемых органов. Исключением являются ганглии, расположенные в непосредственной близости от половых органов. К этим узлам сигналы приходят по длинным преганглионарным волокнам, а передаются к половым органам – по коротким, что обычно бывает характерно для парасимпатического отдела нервной системы.
У млекопитающих парасимпатические ганглии расположены только в области головы и вблизи органов таза. Во всех остальных случаях парасимпатические нейроны лежат в толще и на поверхности легких, сердца и желудочно-кишечного тракта (интрамуральные ганглии).
Парасимпатическими нервами млекопитающих, обслуживающими глазные мышцы и железы, расположенные в области головы, являются глазодвигательный, лицевой и языкоглоточный нервы, выходящие из стволового отдела головного мозга. К органам брюшной и грудной полости парасимпатические нервы идут в составе блуждающего нерва. К органам таза парасимпатические нервы идут из крестцового отдела спинного мозга.
Афферентные нервные волокна входят в состав крупных периферических нервов. В спинной мозг они входят через спинальные ганглии, однако большая их часть попадает в ЦНС в составе блуждающего нерва.
Симпатические постганглионарные нейроны обладают широкой зоной иннервации. Эффект их действия выражается в торможении активности многих внутренних органов и стимуляции сердечной деятельности. Парасимпатическая система действует более локально и оказывает в основном регулирующее действие на функции органов – мишеней. У многих животных в одном и том же органе превалирует либо симпатическая, либо парасимпатическая система. Для низших позвоночных характерно управление многими органами с помощью только одной из систем.
Анатомические преобразования спинного мозга
в ряду хордовых животных.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 |
