

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Пищеварение осуществляется вакуолями. У большинства простейших имеется цитостом (ротовое отверстие), у некоторых - сосательные щупальца. Имеются выделительные и дыхательные вакуоли. Органоидами передвижения могут служить временно образующиеся выступы тела - псевдоподии, жгутики или реснички. В основании органов передвижения находятся цилиндрические базальные тельца, которые происходят из органоида - центриоли, играющего роль в регуляции митотического деления ядра, движения жгутиков (рис. 1).
Размножаются простейшие бесполым путем, посредством деления на две особи или путем множественного деления на большее число особей — шизогонии. В ряде случаев к бесполому размножению присоединяется и половой процесс, при котором происходит или полное слияние клеток и их ядер или временное частичное слияние клеток, сопровождающееся обменом ядерного вещества (конъюгация). Основная форма находящаяся на стадии развития, во время которой осуществляется функция питания роста и бесполого размножения называется вегетативной или трофозоитом. При неблагоприятных условиях или в определенные моменты жизненного цикла простейшие - инцистируются, т. е. окружаются защитной оболочкой и впадают в состояние, при котором все жизненные процессы замедляются. В это время животное не двигается и не принимает пищи, и таким образом имеет возможность переносить различные неблагоприятные условия.
На всех пресноводных рыбах СНГ паразитируют представители 6 типов полцарства Protozoa: жгутиконосцы, саркодовые, споровики, микроспоридии, миксоспоридии, ресничные инфузории.
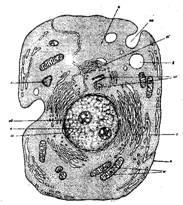
Рис. 1. Схема строения клетки на основании наблюдений в электронном микроскопе:
а - аппарат Голъджи; в - вакуоль; л — лизосома; м - митохондрии; п — плазмалемма; пп - пиноцитозный пузырек; ц — цитоплазма; це - центриолъ; э — эндоплазматическийретикулум; я —ядро; яд —ядрышко; яо - ядерная оболочка.
ОБЩАЯ ХАРАКТЕРИСТИКА МОРФОЛОГИИ И БИОЛОГИИ ЖГУТИКОНОСЦЕВ - ПАРАЗИТОВ РЫБ
Тип Жгутиконосцы - Mastigophora - простейшие, передвигающиеся с помощью жгутиков, расположенных на переднем конце тела. Их бывает 1, 2, 4, 8 и более. Жгутик берет начало в теле животного от базального тела, которое связано своим происхождением с центросомой. Иногда базальное тело связано фибриллами с кинетопластом, имеющим митохондриальную природу. Жгутики свободно свисают или прирастают к телу, образуя ундулирующую мембрану. Тело покрыто пелликулой. Питание или автотрофное (как у растений) или гетеротрофное (как у животных), или сапрофитное. При паразитизме возможно восприятие жидкой пищи всей поверхностью тела. Переваривание идёт в пищеварительных вакуолях, которые образуются вокруг попавшей в цитоплазму пищи. Не переваренные остатки выбрасываются наружу в любом месте заднего конца тела. Размножаются жгутиконосцы продольным делением, реже почкованием или шизогонией. У ряда представителей имеет место половой процесс.
1. Род Trypanosoma (рис. 2).
Тело удлиненное, заостренное на обоих концах, жгутик один. Назальное тельце и кинетопласт находятся на заднем конце тела. Длинная ундулирующая мембрана большей частью переходит на переднем конце тела в свободный жгутик. У рыб трипаносомы паразитируют в кровяном русле. Большая часть жизненного цикла проходит в организме кровососущих пиявок.. В их кишечнике, куда трипаносомы попадают вместе с кровью рыбы, паразиты проходят свои стадии развития принимая различные морфологические формы, после размножения скапливаются в полости хоботка пиявок. Отсюда во время кровососания они снова попадают в кровяное русло рыб. Длина мкм и более. Систематика трипаносом, паразитирующих в рыбах, чрезвычайно запутана.
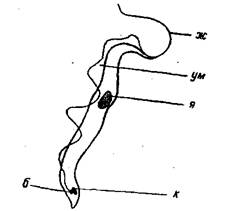
Рис. 2. Схема строения Trypanosoma:
ж — свободная часть жгутика; ум —ундулирующая мембрана; я —ядро; к - кинетопласт; б - базальное тельца
2. Род Cryptobia (рис. 3).
Бесцветные удлиненные жгутиконосцы с двумя жгутиками, один из которых (передний) свободный, а другой (идущий назад) частично прирастает к телу, не образуя, однако, ундулирующей мембраны. Кинетопласт удлинен и слегка изогнут. У рыб встречаются в кишечнике, на жабрах и в кровяном русле. Длина тела от 10 до 27 мкм и более.
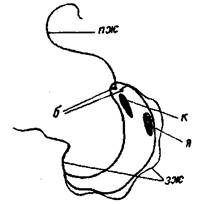
Рис. 3. Схема строения Cryptobia:
пж - передний жгутик; к - кинетопласт; я - ядро; зж - задний жгутик; 6 — базальные зерна.
Возбудитель эктопаразитарного криптобиоза - Cryptobia branchialis (рис. 4).
Вытянутое тело, расширенное на переднем конце и заостренное на заднем. В протоплазме расположены светопреломляющие гранулы. На переднем конце тела и заднем конце - по одному жгутику. Развитие паразита происходит без смены хозяев. Размножение - продольным делением. Вне тела хозяина может плавать в воде 1 - 2 дня. У рыб локализуется на жаберных лепестках. Хозяева: белый и черный амуры, толстолобики, золотой карась, карп, многие сорные и дикие рыбы.
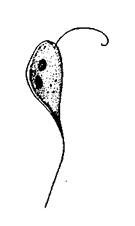
Рис. 4. Cryptobia branchialis
Возбудитель эндопаразитарного криптобиоза - Cryptobia cyprini (рис. 5).
Форма тела варьирует в широких пределах, однако преобладает серповидно изогнутая, расширенная спереди и сужающаяся к заднему концу. Сравнительно длинные жгутики берут начала вблизи крупного, короткого кинетопласта, расположенного обычно ближе к переднему концу тела. Локализация - кровяное русло. Развитие происходит со сменой хозяев - рыб и пиявок, в кишечнике последних паразиты размножаются. Хозяева: карп, сазан, белый амур.

Рис 5. Cryptobia cyprini
Возбудитель костиоза - Costia necatrix (Ichthiohodo necatrbc) (рис. 6).
Тело свальное, с небольшой вогнутостью с брюшной стороны, сбоку -клиновидное; длина 5-20 мкм. Два направленных назад жгутика. Часто встречается еще одна пара жгутиков - результат незавершенного деления. Эти жгутики более короткие и плотно прилегают к телу. Внутри цитоплазмы - маленькое круглое ядро и расположенная поблизости от него сократительная вакуоль. На брюшной стороне имеется продольно изогнутый желобок, заканчивающийся ротовым отверстием. Костию обычно наблюдают в прикрепленном виде на жабрах и коже рыб. Плавает а воде плохо и если не находит хозяина то через 30 мин - 1 час инцистируется. Размножается продольным делением. Питается эпителиальными клетками. Хозяева: практически все выращиваемые рыбы, особенно лососевые, сиговые.

Рис. 6. Costia necatrix
Возбудитель гексамитоза — Hexamita salmonis (Hexamita truttae) (рис. 7).
Грушевидной формы. Длина ,5 мкм. Ширина 3-6 мкм. Имеет 2 ядра, 2 парабазальных тела, расположенных на переднем конце тела. Паразит имеет 4 пары жгутиков, из них 3 пары отходят от переднего конца, а 1 пара, начинаясь с переднего конца, проходит внутри цитоплазмы вдоль всего тела и только на заднем конце выходит наружу. Паразит, как правило, не
прикрепляется к стенке кишечника или желчного пузыря, а плавает в их содержимом. Размножается делением. Заражение происходит путем заглатывания его цист, которые попадают в воду вместе с испражнениями рыб. Хозяева: многие виды лососевых.
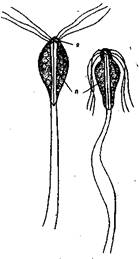
Рис. 7. Hexamita salmon is:
я - ядро; п — парабазальное тело.
II. ТИП СПОРОВИКИ -APICOMPLEXA
Из класса кокцидий в рыбах СНГ паразитируют только представители семейства Eimeridae. Это кокцидий, у которых при спорогонии образуется 4 спороцисты с 2 спорозоитами в каждой из них. У рыб встречаются представители родов - Eimeria и Goussia характеризующиеся наличием овальной или круглой ооцисты. Систематика кокцидий основана на размерах и строении ооцисты. У представителей рода Eimeria на одном конце споры имеется тело Штида, у видов рода Goussia его нет. Паразитируют в эпителиальных клетках кишечника, печени, почек, гонад и других органов рыб. Это внутриклеточные паразиты. Общий ход развития Eimeria следующий (рис. 8). При попадании в рыбу спорозоиты покидают спороцисту, внедряются в клетки органов и растут, превращаясь в округлую или овальную клетку (шизонт) с пузыревидным ядром, которое начинает многократно делиться. Шизонт распадается на большое количество червеобразных мерозоитов (шизогония). Мерозоиты проникают в клетки, растут, превращаясь в шизонта, таким образом происходит усиленное самозаражение хозяина. Стадия шизогонии соответствует острой форме зболевания. После образования нескольких бесполых поколений следует половой процесс и спорообразование. Часть мерозоитов при этом превращается в макрогаметы, которые в свою очередь делятся на большое число микрогамет, снабженных 2 жгутиками. При слиянии макро - и микрогамет образуется зигота, которая превращается в ооцисту, одетую плотной оболочкой. Половое размножение - это хроническое течение болезни. Ядро ооцисты делится на 4 дочерние ядра, вокруг которых обособляются небольшие участки цитоплазмы. Образуются споробласты, одеваются оболочкой и превращаются в спороцисты, внутри которых образуется 2 спорозоита При дифференцировке спороцист, спорозоитов часть цитоплазмы не участвует в этом процессе и остается в виде остаточного тела. Ооцисты выводятся из тела рыбы и могут длительное время находиться в состоянии покоя до нового заражения рыбы.
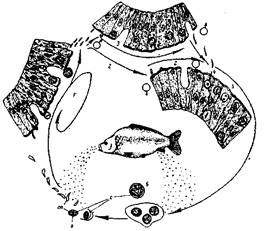
Рис. 8. Цикл развития Eimeria сагреlli (Goussia carpelll):

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 |
