
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Во время откладки яиц были пойманы 6 самок болотной черепахи, у которых были измерены следующие морфометрические показатели:
L. car. – длина карапакса от заднего края загривкового щитка до края надхвостового щитка; Lt. car. – наибольшая ширина карапакса; L. cd. – длина хвоста от переднего края клоакальной щели до кончика.
Возраст животных определялся по числу годовых колец на панцире.
Производился подсчет отложенных и разоренных кладок, числа яиц в них, велись наблюдения за поведением животных. Использованы также отдельные сведения, полученные сотрудниками кафедры биологии Тамбовского госуниверситета и Хоперского заповедника в предыдущие годы.
В апреле черепахи возобновляют активность после зимовки. В апреле – мае происходит спаривание. К концу мая – началу июня самки готовы к откладке яиц. Для этого они выходят на сушу, иногда уходя довольно далеко от водоема. Это явление начинается в вечернее время, около 16–17 ч. и продолжается до ночи. Так, мы находили особей, откладывающих яйца в 21 час.
Для откладки яиц выбираются преимущественно возвышенные места с песчаным грунтом. Самка задними ногами вырывает ямку. Приготовление ямки, по нашим наблюдениям, занимает до 30 минут. Согласно литературным данным (Лукина, 1976), этот процесс продолжается от 1 до 2 часов. Чтобы размягчить почву, животному приходится неоднократно увлажнять ее водой из мочевого пузыря.
Когда ямка готова, самка приступает к откладке. Яйца аккуратно укладываются на дно ямки с интервалами в 1–2 минуты. По литературным данным (Лукина, 1976), эти промежутки более длительные (3–5 минут). После откладывания последнего яйца ямка засыпается и утрамбовывается.
Точное число яиц в кладках установить не удалось. Во время самой кладки животных не беспокоили, а на следующий день все отмеченные кладки оказались разоренными хищниками. В семи разоренных кладках (2007–2008 гг.) по остаткам скорлупы удалось подсчитать примерное число яиц: от 5 до 11 штук (в среднем – 7.14). Вероятно, эти данные несколько занижены, так как некоторые хищники поедают яйца со скорлупой или утаскивают их в другое место. Согласно литературным данным, болотные черепахи в Украинских Карпатах откладывают 6–9, редко 12 яиц (Щербак, Щербань, 1980), Ростовской области и Восточном Приазовье – 5–19 (Лукина, 1976; Гуськов и др., 1983), Волжско-Камском крае – 5–20 (Гаранин, 1983). Отмечается, что плодовитость самок зависит от их возраста (Лукина, 1976).
Размер и возраст самок из нашего материала, участвующих в размножении, приводятся в табл. 1. Самая молодая самка, участвующая в размножении, имела возраст 7 лет. В Украинских Карпатах самки черепах достигают половой зрелости в возрасте 9–10 лет (Щербак, Щербань, 1980), а в южной популяции Восточного Приазовья – 5 лет (Лукина, 1976). Наши данные, возможно, свидетельствуют о более позднем созревании животных из северных популяций.
Таблица 1
Размеры (мм) и возраст (лет) самок болотной черепахи,
участвующих в размножении
№ | L. car. | Lt. car. | L. cd. | Возраст |
1 | 115.5 | 96.1 | 55.4 | 7 |
2 | 171.3 | 143.1 | 88.4 | 10 |
3 | 192.2 | 148.1 | 83.9 | 11 |
4 | 175.9 | 145.6 | 78.8 | 11 |
5 | 152.1 | 123.3 | 89.5 | 9 |
6 | 196.1 | 160.5 | 99.3 | 14 |
После окончания кладки самка не заботится о потомстве и возвращается в водоем.
Инкубационный период – 70–100 суток; по его окончании в августе – октябре из яиц выходят детеныши, которые обычно остаются зимовать в почве до весны будущего года (Ананьева и др., 1998). В некоторых местах, напротив, наблюдалось перемещение молодых черепах в зимовочный водоем в конце лета (Гуськов и др., 1983).
Размеры сеголеток варьируют от 20 до 28 мм (Лукина, 1976; Банников и др., 1977; Щербак, Щербань, 1980; Ананьева и др., 1998). В нашем материале есть два детеныша, пойманных в июне, то есть вскоре после выхода с первой зимовки. Они имели длину карапакса 33.6 и 33.9 мм.
В хоперской популяции, находящейся вблизи северной границы ареала, самки черепах, по всей видимости, делают только одну кладку за сезон. В более теплых климатических условиях число кладок за сезон может достигать трех (Банников, 1951).
Я искренне благодарна , и (Хоперский государственный заповедник) за помощь в организации полевого исследования, , и (Тамбовский государственный университет) – за помощь в сборе и обработке материала.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (грант № ).
Литература
1. , , Орлов и пресмыкающиеся. Энциклопедия природы России. М.: ABF, 19с.
2. , , Барабанов пресмыкающихся Северной Евразии (таксономическое разнообразие, географическое распространение и природоохранный статус). С.-Пб.: Зоологический институт, 20с.
3. Банников к познанию биологии кавказских черепах // Ученые записки Московского городского педагогического института. Т. 18. Вып.С. 129–167.
4. , , Щербак земноводных и пресмыкающихся фауны СССР. М.: Просвещение, 19с.
5. Гаранин и пресмыкающиеся Волжско-Камского края. М.: Наука, 19с.
6. , , Конева земноводных и пресмыкающихся Ростовской области. Ростов-на-Дону: Изд-во Ростовского ун-та, 19с.
7. Лукина размножения болотной черепахи в Восточном Приазовье // Герпетология. Краснодар, 1976. С. 78–80.
8. , Щербань и пресмыкающиеся Украинских Карпат. Киев: Наукова думка, 19с.
Опыт изучения маркировочной активности обыкновенного бобра (саstor fiber l.) в весенний период
, ,
ТГУ им.
Институт естествознания
Экологический научно-образовательный центр
Актуальность данной работы заключается в необходимости изучения детерминант формирования пространственной структуры популяции (Шварц, 1967) для разработки экологических основ управления размножением и пространственным размещением особей с помощью внесения обонятельных сигналов (Muller-Schwarze, Heckman, 1980).
Наиболее востребованными являются результаты, полученные в весенний период времени когда происходит послепаводковое восстановление сети поселений и начинается новый цикл оо - и сперматогенеза животных (Иванова, 1961; Войняк, 1971; Торгун, 1974).
Цель: изучение пространственных и динамических аспектов химической коммуникации бобра в поселениях руслового типа.
Задачи:
ü изучить динамику интенсивности мечения в весенний период;
ü определить приуроченность элементов запаховой коммуникации к пространственно-функциональным зонам поселений;
ü выяснить распределение меток различного типа по инфраструктурным элементам населяемой территории.
Методы:
Обследование осуществлялось на байдарке, с обязательным выходом у любого проявления жизнедеятельности бобров. При этом не допускалось оставление посторонних предметов и изменения облика берегового ландшафта; обувь мылась при каждом выходе. Высадка на берег осуществлялась в стороне от следов наземной активности зверей. Пороговые значения обонятельной чувствительности исследователя равны 3% водного раствора фенола, запах которого сходен с запахом свежей бобровой «струи» (Hinze, 1950). Виды наземной активности бобров, связанные с изучением хемокоммуникации, подразделялись на:
Сигнальный холмик (СХ) – специально сооруженный бобром холмик, используемый в качестве субстрата для ольфакторного мечения территории
Тропа-вылаз (Т-В) – тропа, перемещение по которой не было переходом в соседний водоем или путем к корму.
Тропа кормовая (ТК) – тропа, используемая для выхода к корму.
Тропа-переход (Т-П) – тропа, используемая для перехода в соседний водоем.
Площадка активного мечения (ПАМ) – участок берега, как правило, лишенный растительности с несколькими одиночными метками, располагающимися не далее 1 м друг от друга. ПАМ может быть без сигнальных холмиков, но иметь улавливаемый запах и (или) цвет "струи" в нескольких ее частях
Вылаз с запахом (ВСЗ) – участок берега (вылаза), на котором регистрируется запах бобровой «струи».
Индифферентный участок – участок с запахом бобровой «струи», находящийся вне троп.
Наличие запаха определялось при максимальном приближении к маркировочному субстрату. При необходимости брались пробы материала (во избежание оставления собственного запаха для этого использовались чистое лезвие ножа или одноразовая тонкая палочка). Нахождение всех следов жизнедеятельности отмечалось на смехе русла реки, анализ их размещения на первом этапе предполагал выделение в пределах поселений зон с различным обилием следов наземной активности бобров. На втором этапе оценивалась приуроченность выделенных типов проявлений жизнедеятельности к пространственным зонам, интенсивность создания и обновления меток.
Материалы:
ü Сбор материала проводился в период с 30.03.08 по 25.05.08.
ü Протяженность стационара – 12 км, осуществлено 8 экскурсий, общей протяженностью 96 км.
ü Отмечено 932 проявления наземной активности бобра, из них 219 с запахом бобровой «струи».
ü Инфраструктурные элементы изученных поселений включали 65 троп-переходов, 78 – троп к корму и 90 троп-вылазов, являющихся проявлением исследовательского поведения животных.
Результаты
Проведенное исследование позволило установить основные закономерности освоения и мечения населяемого пространства в весенний период. По результатам работы даны практические рекомендации для получения требуемых поведенческих и физиологических эффектов.
На рисунке 1 отражена динамика численности ольфакторных меток. Степень активности мечения выяснялась по сравнению с предыдущим проходом. Активность проявления ольфакторного мечения в весенний период, напрямую связанна с динамикой числа СХ и ВСЗ. На протяжении всего изучаемого периода численность ПАМ оставалась стабильна, что является следствием специфичности данного вида мечения. (1975) и , (1982) отмечали возникновение участков активного мечения на периферийных участках охраняемых территорий, как проявления «встречной» маркировки пространства. По данным -Никифорова и др. (1961), (1962), увеличение числа ПАМ отмечается в осенний период, в послепаводковое время эта величина остается, относительно неизменна. (1954) связывал увеличение маркировочной активности с сокращением кормовой базы, что приводит к увеличению исследовательской реакции и времени патрулирования, при этом бобры чаще пересекают границы чужих территорий и имеют больше возможностей для проявления «встречного» мечения, и, как следствие, сооружения ПАМ.
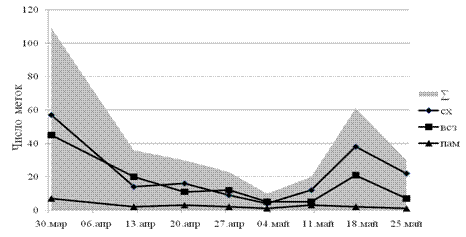
Рисунок 1.Динамика активности ольфакторного мечения
Анализируя встречаемость ольфакторных меток, можно сделать вывод о том, что бобр отдаёт предпочтение СХ (рис. 2), занимающим 2/3 от всех проявлений запаховой коммуникации.
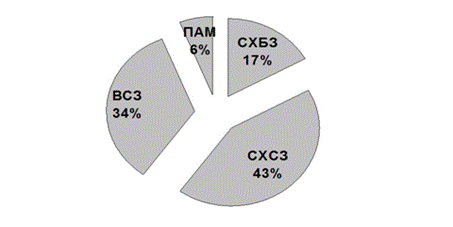
Рисунок 2. Долевое распределение различных типов ольфакторных меток
При сопоставлении динамики изменения уровневого режима р. Ворона с изменением числа всех следов наземной активности, отмечается, что пессимумы маркировочной активности животного приходятся на пик паводка. Анализируя данные с помощью рангового критерия корреляции Спирмена, можно заключить, что между числом следов наземной активности и уровневым режимом воды имеется обратно пропорциональная зависимость средней силы (К = -0,41). Это связанно с тем, что при подъеме уровня воды затопляется пойма реки и значительно сокращаются территории для оставления меток.
При распределении следов наземной активности по зонам поселения (рис. 3)
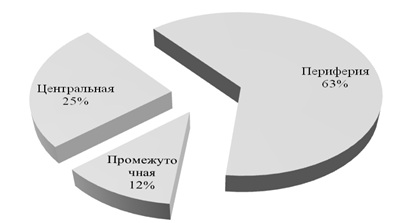
Рисунок 3. Долевое распределение ольфакторных меток по пространственным зонам поселений преобладающее количество сосредоточенно в периферийной и центральной зонах. Эта закономерность объясняется повышенной значимостью данных участков для животного (Емельянов, 2004).
Из рисунка 4 следует, что приоритет в обновлении запаховых меток отдаётся
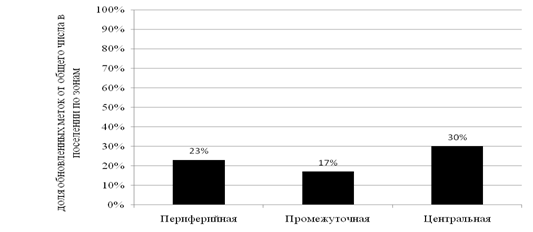
Рисунок 4. Распределение обновлённых ольфакторных меток
центральной, а затем периферийной частям поселения. Это является следствием предпочтения в посещении животными этих участков, в виду их значимости для поддержания целостности пространственной структуры поселений и выполнения функции защиты населяемой территории.
В соотношении числа инфраструктурных элементов поселения (рис. 5) приоритет принадлежит тропам-вылазам. Это связанно с интенсивной исследовательской активностью животного в весенний период.
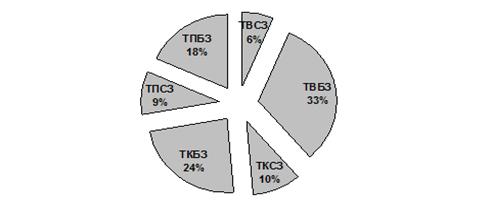
Рисунок 5. Долевое соотношение инфраструктурных элементов в поселениях
Выводы:
ü число СХ и ВСЗ уменьшалось к первой декаде мая, затем имело тенденцию к увеличению;
ü общая интенсивность маркировочной активности закономерно связана с динамикой СХ и ВСЗ ;
ü отмечена обратно пропорциональная зависимость между уровневым режимом реки и интенсивностью мечения;
ü наибольшее число различных меток и их обновляемость отмечены в периферийных зонах, затем следует центральная. Промежуточная зона является мало посещаемой и спорадически маркируемой;
ü среди меток различного типа по численности доминируют СХ (60%);
ü в весенний период времени большая часть троп является проявлением исследовательского поведения животных.
Практические рекомендации: для инициирования требуемых поведенческих и физиологических эффектов необходимо размещать запаховые стимулы в конце марта, или в конце мая, на периферийных частях поселений, выбирая тропы-вылазы имеющие СХ.
Используемая литература:
1. Войняк изменчивость яичников и щитовидной железы у самок речного бобра в течение годового цикла: Автореф. дис. … кандидата биологических наук. – Саратов, 1971. – 16 с.
2. Емельянов экология обыкновенного бобра в бассейне среднего течения р. Ворона // Автореф. дис. … кандидата биологических наук. – Саратов, 2004. – 21 с.
3. Иванова цикличность сперматогенеза речного бобра // Тр. Воронежского гос. зап. – Воронеж, 1961. – Вып. 12. – С. 117-139.
4. Наумов (сигнальные) поля и их значение в жизни млекопитающих // Вестник АН СССР. – 1975. – Вып. 2. – С. 55-62.
5. , Рожнов сфера экспериментальных группировок и маркировочное поведение американских норок // Ферромоны и поведение. – М.: Наука, 1982. – С. 219-237.
6. Торгун и кариометрические исследования коры надпочечников у самцов речных бобров в зависимости от сезона и половой функции // Тр. Воронежского гос. зап. – Вып. 20. – Воронеж, 1974. – С. 24-30.
7. Шварц структура вида // Зоологический журнал. – 1967. – Т. 46. – Вып. 10. – С. .
8. Hinze G. Biber. Korperbau und Lebensweise, Verbreitung und Geschicht. – Berlin, Akademie-Verlag, 1950. – 216 s.
9. Muller-Schwarze D., Heckman S. The social role of marking beaver (Castor canadensis) // J. of Chemical Ecology. – 1980. – Vol. 6. – № 1. – P. 81-95.
Работа выполнена в рамках ИОП по направлению «Живые системы»
Экология и морфология ротана-головешки (Perccottus glenii)
в водоемах степных западин Тамбовской области
, 4 курс,
Институт естествознания ТГУ им.
Ю,
Мичуринский государственный педагогический институт
Ротан (Perccottus glenii Dybowski, 1877) – некрупная рыба, сильно изменившая свой ареал в результате действий человека. Естественный ареал вида расположен на Дальнем Востоке России, в северо-восточном Китае и на севере Северной Кореи. Наиболее обычен в бассейне Амура (Берг, 1949). Неприхотливость ротана позволила ему прижиться в тех местах, куда его выпускал человек.
В результате искусственного расселения ротан в начале XX века появился в естественных водоемах европейской части России (Васильева, 2004). На территории Липецкой области в бассейне Дона он стал отмечаться с середины 80-х годов прошлого века (Сарычев, 2007).
Первые сведения о ротане в Тамбовской области приводит (1986). С этого момента ротан отмечается во всех литературных источниках, посвященных ихтиофауне области (Доклад…, 1997; Скрылева и др, 1999; Скрылева и др., 2002; Соколов и др, 2007).
Данное сообщение посвящено особенностям экологии и морфологии ротана P. glenii в замкнутых водоемах степных западин Тамбовской области.
9–10 июля 2008 года были обследованы 2 водоема естественного происхождения в блюдцеобразных понижениях степных западин на водоразделе бассейнов рек Воронеж и Матыра в Петровском районе. Этим водоемам были присвоены условные названия Большой и Малый в соответствии с их размером. Малое западинное озеро находится в 2.5 км к северу от с. Петровское, имеет глубину до 1.4 м, диаметр 20 м, дно илистое, слой ила 20–30 см, сильно заросло элодеей и рдестами. Обловленный участок – 120 м2. Большое западинное озеро расположено в 4 км к северу от с. Петровское, имеет глубину более 2 м, дно илистое, слой ила у берега 30–50 см, сильно заросшие рогозом и тростником берега. Обловленный участок – 100 м2.
Отлов производился ставными сетями и сетью Кинелева. Собранный материал фиксировался в 4% формалине. В двух водоемах были пойманы 74 особи ротана. Измерение пластических признаков проводились по схеме, общей для семейства окунеобразные, для определения возраста использовалась чешуя (Правдин, 1935). Чешуя для определения возраста бралась в передней части туловища выше боковой линии. Подсчет производился в соответствии со шкалой возрастов (Никольский, 1974). Использована шкала для определения зрелости половых продуктов (I–VI стадии) (Кошелев, 1984).
У добытых рыб измерялись 26 пластических признаков: TL – общая длина тела; l – длина тела до начала лучей хвостового плавника; aD – антедорсальное расстояние; pD – постдорсальное расстояние; lD I – длина основания первого спинного плавника; hD I – высота первого спинного плавника; lD II – длина основания второго спинного плавника; hD II – высота второго спинного плавника; aA – антеональное расстояние; P-V – расстояние от основания грудного плавника до основания брюшного; V-A – расстояние от основания брюшного плавника до основания анального; lP – длина грудного плавника; lV – длина брюшного плавника; lA – длина основания анального плавника; hA – высота анального плавника; H – наибольшая высота тела; h – высота хвостового стебля; pl – длина хвостового стебла; w – толщина хвостового стебля в средней части; W – максимальная толщина тела; c – длина головы; ao – длина рыла; o – диаметр глаза по горизонтали; po – заглазничное расстояние; Нс – высота головы у затылка; hc – высота головы через середину глаза. На основании этих признаков были подсчитаны наиболее важные индексы: TL/l, aD/l, pD/l, lD I/l, hD I/l, lD II/l, hD II/l, aA/l, P-V/l, V-A/l, lP/l, lV/l, lA/l, hA/l, H/l, h/l, pl/l, w/lУ, W/l, c/l, ao/c, o/c, po/c, Нс/c, hc/c.
Питание изучалось методом вскрытия желудочно-кишечного тракта. Для определения водных беспозвоночных использовался «Определитель пресноводных беспозвоночных России и сопредельных территорий» (1995, 1997, 1999).
Статистическая обработка данных проводилась стандартным способом (Лакин, 1990) с использованием программы Microsoft Excel.
При изучении ихтиологического разнообразия водоемов, кроме ротана, никаких других видов рыб обнаружено не было. Хотя, по свидетельствам местных жителей, раньше там обитал серебряный карась Carassius auratus (Linnaeus, 1758).
При обследовании Большого западинного водоема, кроме ротанов обычной темной окраски, были добыты 3 экземпляра с частичной депигментацией покровов. Они составили приблизительно 4% от числа ротанов, пойманных в этом водоеме.

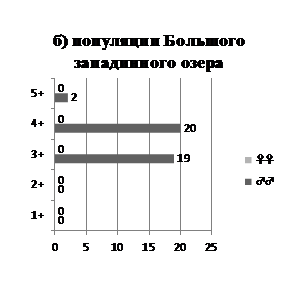
Рис. 1. Половозрастная структура популяций Малого (а) и Большого (б) западинных озер.
Сведения о половозрастной структуре популяций приводятся на рис. 1. В Малом западинном озере численно преобладают самки, среди возрастных групп доминируют особи 2- и 3-летнего возраста. В Большом западинном озере популяция представлена в основном самцами, возраст которых 3–4 года. В этом озере были пойманы две крупные самки (общая длина тела – 25 см), возраст которых не был установлен.
Сравнение мужских и женских особей из Малого водоема (табл. 1) дало следующие результаты. При сравнении абсолютных показателей оказалось, что самцы и самки отличаются по 25 из 26 признаков. Значение TL, l, aD, pD, lD1, hD1 выше у самцов, а lD2, hD2, aA, V-A, lP, lV, lA, hA, H, h, pl, w, W, c, ao, o, po, Нс, hc – у самок. При сравнении индексов различий оказалось значительно меньше, как по числу отличающихся признаков, так и по степени их достоверности. Значение индексов TL/l, V-A/l, lV/l, o/c выше у самок, а lD2/l, lP/l, lA/l, ao/c – у самцов.
Кроме того, имеются отличия в строении урогенитальных сосочков: у самок обычно 2 урогенитальные поры, у самцов – 1. Связано это, прежде всего, со сложным нерестовым поведением и особым способом откладки икры.
Определение стадий зрелости половых продуктов самок дало следующие результаты. У двух самок из Большого западинного озера отмечена IV стадия зрелости половых продуктов, плодовитость – 5400 и 5750 икринок. 15 самок из Малого западинного озера находились на различных стадиях зрелости икры: 2 самки на I стадии, 4 – на II, 7 – на III и 2 – на IV. У двух самок на IV стадии зрелости плодовитость составила 179 и 354 икринок.
В рационе ротана были выявлены представители следующих таксонов.
Ракообразные: Ветвистоусые раки Daphniiformes, Веслоногие раки Calanoida, Ракушковые раки Ostrocoda.
Паукообразные: Акариоморфные клещи Acariformes.
Насекомые: Поденки Ephemeroptera, Стрекозы Odonata, Полужесткокрылые Heteroptera, Двукрылые Diptera, Жесткокрылые Coleoptera.
Рыбы. Так как в водоеме, кроме ротана, других видов рыб обнаружено не было, это свидетельствует о наличии каннибализма.
Водоросли и детрит.
Таблица 1
Сравнительная характеристика размерных показателей ротана из Малого водоема (курсивом выделены достоверные различия при p ≤ 0.05; полужирным – p ≤ 0.01; полужирным курсивом – p ≤ 0.001)
№ | Показатель | ♂♂ (n=10) | ♀♀ (n=15) | t | ||
lim | M ± m | lim | M ± m | |||
1 | TL | 71,0-90,0 | 78,9 ± 2,05 | 60,0-81,0 | 66,8 ± 1,35 | 4,902 |
2 | l | 59,0-76,0 | 65,4 ± 1,85 | 49,5-67,0 | 54,5 ± 1,17 | 4,965 |
3 | aD | 25,0-33,0 | 27,8 ± 0,90 | 21,0-29,0 | 23,5 ± 0,53 | 4,125 |
4 | pD | 17,0-22,0 | 19,3 ± 0,51 | 13,5-18,5 | 15,9 ± 0,34 | 5,495 |
5 | lD1 | 4,0-8,0 | 6,2 ± 0,37 | 3,0-6,0 | 4,8 ± 0,23 | 3,271 |
6 | hD1 | 8,5-13,5 | 11,3 ± 0,50 | 7,5-12,0 | 9,1 ± 0,29 | 3,640 |
7 | lD2 | 10,0-13,0 | 11,0 ± 0,33 | 7,0-11,0 | 8,6 ± 0,27 | 5,631 |
8 | hD2 | 11,5-17,5 | 13,6 ± 0,59 | 9,0-14,0 | 11,4 ± 0,34 | 3,185 |
9 | aA | 35,0-46,0 | 38,9 ± 1,23 | 30,0-41,0 | 32,9 ± 0,79 | 4,090 |
10 | P-V | 3,5-6,0 | 5,0 ± 0,30 | 3,5-6,0 | 4,3 ± 0,19 | 1,895 |
11 | V-A | 15,0-19,0 | 16,6 ± 0,45 | 13,0-18,0 | 14,5 ± 0,36 | 3,626 |
12 | lP | 12,5-15,0 | 14,0 ± 0,31 | 9,0-14,5 | 11,0 ± 0,40 | 5,747 |
13 | lV | 9,0-11,5 | 10,0 ± 0,27 | 8,0-11,0 | 9,0 ± 0,28 | 2,333 |
14 | lA | 7,0-10,0 | 8,9 ± 0,32 | 6,0-9,0 | 7,0 ± 0,22 | 4,979 |
15 | hA | 11,5-15,0 | 13,0 ± 0,37 | 9,0-14,0 | 11,0 ± 0,35 | 3,753 |
16 | H | 13,5-18,5 | 15,8 ± 0,53 | 11,0-15,0 | 13,0 ± 0,32 | 4,516 |
17 | h | 6,0-9,0 | 7,3 ± 0,32 | 5,0-7,0 | 6,1 ± 0,15 | 3,381 |
18 | pl | 17,0-23,0 | 19,0 ± 0,59 | 13,0-19,5 | 15,4 ± 0,45 | 4,842 |
19 | w | 4,0-6,0 | 4,8 ± 0,25 | 3,0-5,0 | 4,0 ± 0,16 | 2,823 |
20 | W | 8,5-12,0 | 10,2 ± 0,34 | 7,0-11,0 | 8,7 ± 0,30 | 3,291 |
21 | c | 20,5-28,0 | 23,8 ± 0,68 | 18,0-24,0 | 19,5 ± 0,44 | 5,173 |
22 | ao | 5,0-7,0 | 5,7 ± 0,23 | 4,0-7,0 | 4,5 ± 0,23 | 3,858 |
23 | o | 5,0-6,0 | 5,4 ± 0,15 | 4,0-5,5 | 4,8 ± 0,12 | 3,381 |
24 | po | 11,0-16,0 | 13,0 ± 0,47 | 9,0-13,5 | 10,6 ± 0,32 | 4,198 |
25 | Hc | 12,0-16,0 | 13,7 ± 0,47 | 10,0-14,0 | 11,4 ± 0,29 | 4,228 |
26 | hc | 8,0-11,0 | 9,5 ± 0,32 | 7,0-8,5 | 7,5 ± 0,15 | 5,420 |
27 | TL/l | 1,18-1,23 | 1,21 ± 0,004 | 1,20-1,25 | 1,23 ± 0,004 | 3,394 |
29 | aD/l | 0,41-0,43 | 0,43 ± 0,003 | 0,40-0,45 | 0,43 ± 0,004 | 1,164 |
30 | pD/l | 0,28-0,31 | 0,29 ± 0,003 | 0,27-0,32 | 0,29 ± 0,003 | 0,006 |
31 | lD1/l | 0,07-0,11 | 0,09 ± 0,004 | 0,06-0,11 | 0,09 ± 0,004 | 1,262 |
32 | hD1/l | 0,14-0,21 | 0,17 ± 0,005 | 0,13-0,18 | 0,17 ± 0,004 | 0,932 |
33 | lD2/l | 0,14-0,18 | 0,17 ± 0,003 | 0,14-0,17 | 0,16 ± 0,002 | 2,928 |
34 | hD2/l | 0,18-0,27 | 0,21 ± 0,008 | 0,17-0,24 | 0,21 ± 0,005 | 0,141 |
35 | aA/l | 0,58-0,61 | 0,60 ± 0,004 | 0,55-0,65 | 0,60 ± 0,007 | 0,884 |
36 | P-V/l | 0,06-0,09 | 0,08 ± 0,003 | 0,06-0,10 | 0,08 ± 0,003 | 0,930 |
37 | V-A/l | 0,24-0,27 | 0,26 ± 0,002 | 0,24-0,29 | 0,27 ± 0,004 | 2,453 |
38 | lP/l | 0,19-0,24 | 0,21 ± 0,004 | 0,18-0,23 | 0,20 ± 0,004 | 2,360 |
39 | lV/l | 0,14-0,18 | 0,15 ± 0,003 | 0,14-0,19 | 0,17 ± 0,004 | 2,450 |
40 | lA/l | 0,12-0,15 | 0,14 ± 0,003 | 0,12-0,14 | 0,13 ± 0,002 | 2,760 |
41 | hA/l | 0,18-0,22 | 0,20 ± 0,004 | 0,18-0,23 | 0,20 ± 0,004 | 0,406 |
42 | H/l | 0,22-0,26 | 0,24 ± 0,004 | 0,21-0,27 | 0,24 ± 0,004 | 0,159 |
43 | h/l | 0,10-0,12 | 0,11 ± 0,002 | 0,10-0,13 | 0,11 ± 0,002 | 0,676 |
44 | pl/l | 0,26-0,31 | 0,29 ± 0,004 | 0,26-0,32 | 0,28 ± 0,004 | 1,385 |
45 | w/l | 0,06-0,08 | 0,07 ± 0,002 | 0,06-0,09 | 0,07 ± 0,002 | 0,291 |
46 | W/l | 0,14-0,17 | 0,15 ± 0,003 | 0,14-0,20 | 0,16 ± 0,004 | 1,095 |
47 | c/l | 0,34-0,37 | 0,36 ± 0,003 | 0,35-0,37 | 0,36 ± 0,002 | 1,281 |
48 | ao/c | 0,21-0,29 | 0,24 ± 0,007 | 0,20-0,27 | 0,22 ± 0,005 | 2,525 |
49 | o/c | 0,21-0,24 | 0,23 ± 0,004 | 0,22-0,27 | 0,25 ± 0,005 | 2,832 |
50 | po/c | 0,50-0,57 | 0,55 ± 0,006 | 0,49-0,58 | 0,54 ± 0,008 | 0,594 |
51 | Hc/c | 0,53-0,62 | 0,58 ± 0,009 | 0,55-0,62 | 0,58 ± 0,006 | 0,406 |
52 | hc/c | 0,35-0,43 | 0,39 ± 0,008 | 0,37-0,43 | 0,39 ± 0,005 | 0,569 |
Таким образом, изученный нами рацион ротана в водоемах степных западин Тамбовской области близок к таковому в популяциях вида европейской части России (http://www. *****) и Западной Сибири (Дубчак, 2005). Единственной характерной особенностью является наличие мелких ракообразных в желудках рыб размером 11–16 см из Большого водоема, в то время как в научной литературе пишется, что эта пища свойственна ротанам, не превышающим 5–6 см. Это может быть связано с переходом ротана на питание бентосными беспозвоночными из-за отсутствия другого подходящего корма. Указанный выше каннибализм был отмечен у особей всех изученных возрастных групп в обоих водоемах. Это явление не уникально и было ранее описано у ротана, обитающего в небольших замкнутых водоемах.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 |
