


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Отдел Eumycota (истинные грибы). По морфологии различают мицелярные и дрожжевые формы грибов.
Мицелиальные грибы
Морфология. Тело мицелиальных грибов представляет собой систему из длинных ветвящихся нитей – гифов. Гифы образуют одно - или многоклеточное вегетативное тело гриба – таллом (грибница). Всю совокупность гиф грибного таллома называют мицелием. Способность к образованию мицелия – один из отличительных признаков истинных грибов (Eumycota). Иногда гифы сливаются своими оболочками, образуя псевдопаренхиму. На отдельные клетки гифы разделяют септы (перегородки). У низших грибов гифы не имеют перегородок, их клетки многоядерные. Мицелий низших грибов называется ценоцитным (несептированным). У высших грибов мицелий септированный. Различают вегетативный и спороносящий (репродуктивный) мицелий. Последний образует специализированные гифы, несущие репродуктивные структуры – спорофоры. Вегетативный мицелий большинства видов не окрашен, поэтому молодые колонии таких грибов белые или сероватые. По мере развития органов плодоношения колонии приобретают зеленую, желтую, коричневую, красную, малиновую или черную окраску. У некоторых грибов пигмент диффундирует в среду. На плотных средах мицелиальные грибы образуют мягкие, не врастающие в субстрат, округлые или широко распространяющиеся по поверхности, пушистые, нитевидые, паутинообразные, ватоподобные или порошкообразные колонии.
Диаметр гиф грибов колеблется от 5 до 50 мкм. Клеточная стенка грибковой клетки содержит полисахариды, преимущественно хитин, а также хитинозан, глюканы, маннаны (пептидогликан отсутствует). С возрастом клеточная стенка утолщается, в ней откладываются высокомолекулярные липиды, минеральные соли, пигменты. Фимбрий и жгутиков грибковая клетка не имеет. Мицелиальные грибы – неподвижные организмы. У некоторых грибов образуются подвижные за счет сложно устроенных жгутиков зооспоры. Цитоплазматическая мембрана в качестве основных стероидов включает эргостерин и зимэстерол. В цитоплазме содержится большое количество рибосом, имеются митохондрии, лизосомы, вакуоли, включения гликогена, волютина (крахмал отсутствует), развитая эндоплазматическая сеть. В клетках грибов четко дифференцировано ядро, окруженное ядерной мембраной. Несептированный мицелий грибов содержит несколько ядер.
Тип питания грибов хемоорганогетрофный, голофитный. Грибы – аэробные организмы, реже факультативные анаэробы или микроаэрофилы. Большинство грибов мезофилы, но встречаются психрофилы и термофилы. Имеются ацидофилы, алкалофилы, галотолерантные и галофильные виды.
Способы размножения грибов разнообразны. У них возможно вегетативное, бесполое и половое размножение. Вегетативное размножение осуществляется отдельными участками мицелия, т. е. без образования специализированных органов размножения.
Бесполое размножение происходит при помощи спор. Споры, развивающиеся эндогенно (внутри терминального увеличенного конца гифы), называют эндоспорами. Они заключены в специализированные мешкообразные структуры – спорангии, которые формируются на свободных концах плодоносящих гиф – спорангиеносцах (спорангиофоры). Подобный тип спорообразования характерен для грибов рода Mucor. Если споры располагаются на поверхности спороносящего органа, то их называют экзоспоры или конидии. Число, форма и размеры конидий у разных видов существенно варьируют. У некоторых видов репродуктивные структуры имеют особую организацию. Так, у видов рода Aspergillus репродуктивные структуры (конидиефоры, или конидиеносцы) заканчиваются терминальными пузырьками или головками, на которых располагаются выросты, называемые стеригмами. На стеригмах образуются одноклеточные конидии, располагающиеся в виде цепочки. У видов рода Penicillium конидиеносцы двукратно ветвятся (рис. 29). Иногда конидии могут развиваться при фрагментации гифального тяжа, их обозначают как артроконидии.
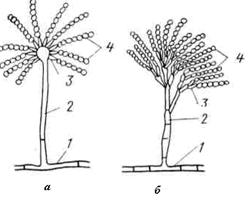
Рис. 29. Конидиеносцы у грибов рода Aspergillus (а) и Penicillium (б);
1 — вегетативный мицелий, 2 — конидиеносец, 3 — стеригмы, 4 — конидии
Споры грибов воздушными течениями могут переноситься на большие расстояния. Так, споры возбудителя стеблевой ржавчины пшеницы переносились на 1000 км от источника их массового развития.
Половое размножение заключается в слиянии мужских и женских гамет, в результате чего образуется зигота. У низших грибов половой процесс состоит в конъюгации одинаковых или разных по размерам гамет (изо - или гетерогамия) или имеет место оогамный половой процесс. В последнем случае развиваются женские (оогонии) и мужские (антеридии) половые органы. В оогониях развивается несколько яйцеклеток или одна из них. Оплодотворение яйцеклетки происходит или сперматозоидами, или выростом антеридия, переливающим в оогоний свое содержимое. У низших грибов половой продукт (ооспора) прорастает в спорангий со спорами.
При половом размножении зигомицетов между гифами одного мицелия (у гомоталличных видов) или разных мицелиев (у гетероталличных видов) образуются короткие поперечные выросты, отделяющие на концах многоядерные клетки, называемые гаметангиями (рис. 30). Они сливаются и образуют ценозиготу, которая увеличивается в размерах и превращается в толстостенную зигоспору. После периода покоя зигоспора прорастает с образованием зародышевого спорангия. При этом имеет место мейоз. Многоядерная цитоплазма спорангия распадается на множество спорангиеспор, каждая из которых, освободившись при разрыве спорангия, в соответствующих условиях может прорасти в мицелий. Такой половой процесс называют гаметангиогамия.
У сумчатых грибов фаза полового размножения начинается с образования аскогонов. Аскогон имеет особую трубочку – трихогину, в которую попадают мужские ядра. Эти ядра через трихогину переходят из антеридиев в аскогон, где происходит плазмогамия. Затем из аскогона вырастают так называемые аскогенные гифы, клетки которых содержат по одному мужскому и одному женскому ядру (дикариотические гифы). Эти ядра одновременно делятся. Слиянию ядер дикариона предшествует специальное деление клетки с образованием крючка: кончик гифы крючкообразно искривляется, происходит сопряженное деление обоих ядер и верхняя пара дочерних ядер отделяется поперечными перегородками как от клетки, лежащей ниже изгиба, так и от крючка. Крючок сливается с клеткой, находящейся ниже изгиба, и в результате снова образуется клетка с двумя ядрами. Верхняя клетка в дальнейшем становится аском (сумкой). В ней происходит слияние обоих ядер. Образовавшееся первичное ядро аска (диплоидное) претерпевает три деления, одно из которых мейотическое; в результате получается восемь дочерних ядер и, в конечном счете, восемь аскоспор. Число делений может быть меньшим (в этом случае образуется 4 споры) или большим (число спор иногда превышает 1000). У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших в специальных вместилищах – плодовых телах. Различают следующие типы плодовых тел: клейстотеци – полностью замкнутые плодовые тела, характерны для плектомицетов; перитеции – полузамкнутые, обычно кувшиновидные плодовые тела с отверстием на вершине, типичны для пиреномицетов; апотеции – открытые, обычно чашевидные плодовые тела, характерны для дискомицетов.
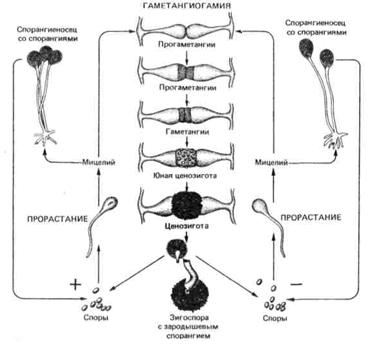
Рис. 30. Цикл развития Rhizopus nigricans
Экология грибов. Грибы широко распространены в природе на самых различных субстратах. Обширна группа почвенных грибов. Физические особенности почвы (пористое пространство между минеральными частицами, недостаток влаги) обуславливают преимущества для мицелиальных форм. Высокая активность грибов в почве объясняется их устойчивостью к изменяющимся условиям среды. Имея неодинаковый оптимум рН для развития, грибы хорошо переносят любые условия кислотности и поэтому встречаются в кислых и щелочных почвах. Большинство почвенных грибов сапрофиты, играют важную роль в процессах почвообразования. Грибы, обладая мощным ферментативным аппаратом, участвуют в разложении таких трудных для разложения соединений, как гумус, лигнин, хитин, целлюлоза. Грибы активно участвуют в превращениях соединений азота, способствуют улучшению структуры почвы, агрегируя почвенные частицы.
Грибы – микоризообразователи вступают в симбиотические отношения с наземными растениями, образуя с их корнями микоризу, участвуют в процессах роста древостоев, а также в их защите от фитопатогенных грибов. К такому симбиозу склонны представители классов Zygomycetes, Ascomycetes, но главным образом Basidiomycetes (агариковые и болетовые).
Для водной среды характерны представители классов Chytridiomycetes и Oomycetes. В основном это паразиты водных беспозвоночных животных или водорослей.
Среди грибов имеются паразиты растений и животных. Большой урон сельскому хозяйству наносят фитопатогенные грибы. Энтомопатогенные грибы, например, энтомофторовые (порядок Entomophthorales), несовершенные грибы родов Beauveria, Paecilomyces, Metarhizium, Verticillium и другие, являются возбудителями микозов у насекомых, их используют для создания микоинсектицидных препаратов для регулирования численности насекомых – вредителей.
Кератинофильные плесневые грибы (представители родов Trichophyton, Microsporum, Epidermophyton) вызывают дерматомикозы у человека. Они выделяют кератиназу – фермент, разлагающий кератин эпидермиса, волос и ногтей.
Существуют специфические экологические группы грибов, которые вызывают деструкцию различных промышленных материалов: бумаги, тканей, кожи, резины, пластмассы, бетона, камня, мрамора и т. д.
В процессе жизнедеятельности грибов выделяются различные физиологически активные вещества – ферменты, органические кислоты, витамины, антибиотики, токсины.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 |
