


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Запасные вещества – полисахариды, липиды, белки, полипептиды, полифосфаты, отложения серы, углеводороды. Полисахариды (гликоген, крахмал, гранулеза – крахмалоподобное вещество) при неблагоприятных условиях используется как источник углерода и энергии. Липиды откладываются в виде гранул, резко преломляющих свет; при неблагоприятных условиях используется как источник углерода и энергии. Полифосфаты содержатся в гранулах (волютиновые зерна) и используются как источник фосфора. У серных бактерий сера откладывается в виде глобул как промежуточный продукт, а при исчерпании из среды H2S бактерии окисляют внутриклеточную серу. Белки – у цианобактерий белок цианофицин используется как запасной источник энергии, азота, углерода. Белковые включения Bacillus thuringiensis представляют собой токсин (δ-эндотоксин), активный против личинок чешуекрылых (рис. 11).


Рис. 11. Кристалл эндотоксина штамма B. thuringiensis ssp. dendrolimus К 30-3
(увеличение 40 000 раз)
Внутрицитоплазматические мембраны
Внутрицитоплазматические мембраны являются производными ЦПМ, возникшие в результате ее разрастания и глубокой инвагинации; увеличивают поверхность мембраны.
Фотосинтетические мембраны имеются у фототрофных прокариот. На них локализован фотосинтетический аппарат. Морфология фотосинтетических мембран разнообразна. Различают везикулярные формы (имеют форму пузырьков), ламеллы (имеют форму пластинок, уложенных в форме стопок), тилакоиды (в виде уплощенных замкнутых дисков) (рис. 12).
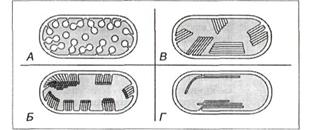
Рис. 12. Строение внутрицитоплазматических фотосинтезирующих мембран у фототрофных
бактерий: А – Мембраны везикулярного типа, присутствующие у Rhodospirillum rubrum, Chromatium
vinosum и др. видов; Б – Трубчатые мембраны у Thiocapsa pfennigii; В – Плоские тилакоидоподобные
мембраны в правильных стопках у Rhodospirillium molischianum; Г – Крупные тилакоиды, частично в
стопках или свободнолежащие у Rhodopseudomonas palustris.
Мезосомы – это локальные впячивания ЦПМ, связаны с клеточным делением и процессом спорообразования.
Система внутрицитоплазматических мембран у хемолитотрофных бактерий (нитрифицирующих, азотфиксирующих, метанотрофных). У этих бактерий высокая активность дыхания.
Клеточная оболочка
Цитоплазматическая мембрана
Химический состав. ЦПМ – это белково-липидный комплекс. Белки составляют 50 – 75 %. Бактериальные ЦПМ относятся к мембранам с наиболее высоким содержанием белка, так как у бактерий нет митохондрий, хлоропластов и все энергетические процессы связаны с ЦПМ. Липиды составляют 15–45 %. Главным липидным компонентом ЦПМ бактерий являются фосфолипиды. У многих бактерий в составе ЦПМ обнаруживаются гликолипиды, аминолипиды, гопаноиды (производные тритерпеноидов), у фототрофных бактерий – каротиноиды. Углеводы составляют от 1 до 16–19 %.
Структура мембран. У бактерий ЦПМ имеет бислойную структуру, сформированную фосфолипидами, связанными с белками. Интегральные мембранные белки пронизывают липидный бислой. Периферические мембранные белки более или менее свободно связаны с одной из поверхностей мембраны и входят в состав асимметрично организованных функциональных комплексов (рис. 13).
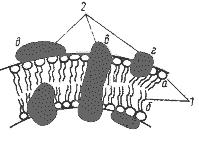
Рис. 13. Модель строения элементарной биологической мембраны:
1 — молекулы липидов: а — гидрофильная "голова"; б — гидрофобный "хвост";
2 — молекулы белков: в — интегральная; г — периферическая; д — поверхностная.
Мембранные белки делят на структурные и функциональные. Функциональные белки участвуют в синтетических и окислительно-восстановительных реакциях, выступают в качестве специфических рецепторов, ответственных за распознавание. Интегральные белки играют роль переносчиков (пермеазы). Липидный бислой также содержит хиноны – переносчики электронов. Компоненты мембраны способны свободно перемещаться в плоскости мембраны.
Функции ЦПМ:
1. Выполняет барьерную функцию, т. е. обеспечивает сохранение существенных различий между клеточным содержимым и окружающей средой.
2. Обеспечивает избирательный транспорт веществ.
3. Выполняет роль осмотического барьера. Благодаря мембране устанавливается разница в концентрации ионов внутри клетки и во внеклеточном пространстве.
4. Метаболическая функция. ЦПМ играет важную роль в превращениях клеточной энергии, в ней локализованы дыхательные цепи, фотосинтетические электрон-траспортные цепи.
5. Сенсорная функция. ЦПМ воспринимает внешние сигналы, что позволяет клетке быстро отвечать на изменения, происходящие во внешней среде
6. Репродуктивная функция (участвует в процессе деления клетки).
Клеточная стенка
Клеточная стенка – важный и обязательный структурный элемент большинства прокариот – обладает определенной ригидностью, жесткостью, в то же время пластична, придает клеткам определенную форму. Клеточная стенка отсутствует у микоплазм и L-форм.
По строению и химическому составу клеточная стенка прокариот отличается от таковой эукариот. В ее состав входит пептидогликан (муреин) – специфический гетерополимер прокариот.
Пептидогликан построен из чередующихся остатков N-ацетил-глюкозамина и N-ацетилмурамовой кислоты, соединенных между собой β-1,4-гликозидными связями. К N-ацетилмурамовой кислоте присоединен короткий пептидный хвост, состоящий из 4-5 аминокислот (глутаминовая, глицин, аланин, диаминопимелиновая (двухосновная – у актиномицетов). Аминокислоты находятся в Д-форме (неприродная конфигурация). Для пептидогликана характерно высокое содержание аминокислот с двумя аминогруппами, что имеет значение для его пространственной организации (обе аминогруппы могут участвовать в образовании пептидных связей) (рис. 14).
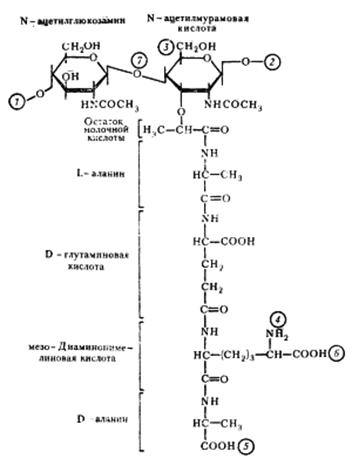
Рис. 14. Структура повторяющейся единицы пептидогликана клеточной стенки эубактерий:
1, 2— места полимеризации гликанового остова молекулы: 3 — место присоединения с помощью фосфодиэфирной связи молекулы тейхоевой кислоты в клеточной стенке грамположительных эубактерий; 4, 5 — места, по которым происходит связывание между гликановыми цепями с помощью пептидных связей; 6 — место ковалентного связывания (пептидная связь) с липопротеином наружной мембраны у грамотрицательных эубактерий; 7 — место действия лизоцима
Молекула пептидогликана представляет собой правильную сеть из параллельно расположенных полисахаридных цепей, соединенных короткими пептидными хвостами. Обладает прочностью и упругостью.
Со структурой и составом клеточной стенки бактерий связаны их тинкториальные признаки, т. е. способность вступать в реакции с красителями и окрашиваться определенным образом.
В 1884 году Грам Ханс Христиан, датский бактериолог, фармаколог и врач, предложил базовый метод дифференциального окрашивания бактерий. Метод заключается в том, что фиксированный препарат бактерий сначала окрашивают генциан фиолетовым, затем обрабатывают раствором Люголя, содержащим йод. Краситель образует с йодом комплекс, умеренно растворимый в этаноле, но не растворимый в воде, что позволяет дифференцировать бактерии на грамположительные – их клетки удерживают комплекс йода с красителем и остаются темно-фиолетовыми – и грамотрицательные, которые обесцвечиваются этанолом. Комплекс генциан фиолетового с йодом локализован, по-видимому, на плазматической мембране. Положительная окраска бактерий по Граму обусловлена тем, что клеточная стенка данных бактерий препятствует экстракции этого комплекса, что связано со значительной толщиной пептидо-гликанового слоя. У грамотрицательных бактерий клеточная стенка не обладает таким свойством.
Окраска по Граму – признак видоспецифичный, используется для определения вида бактерий. Окрашивают только односуточные, молодые культуры, так как способность удерживать окрашенный комплекс зависит от физиологического состояния бактерий.
Клеточная стенка грамположительных бактерий
Клеточная стенка грамположительных бактерий имеет толщину 20 – 80 нм, что соответствует примерно 40 молекулам пептидогликана. Под электронным микроскопом выглядит как гомогенный электронно-плотный слой, прилегающий к ЦПМ (рис. 15).
А 2 3 4 Б
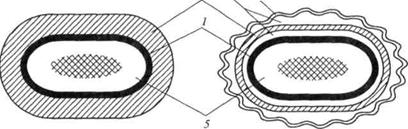
Рис. 15. Клеточная стенка грамположительных (А) и грамотрицательных (Б) эубактерий:
1 – цитоплазматическая мембрана, 2 – пептидогликан, 3 – периплазматическое пространство, 4 – наружная мембрана, 5 – цитоплазма, в центре которой расположена ДНК
У грамположительных бактерий основным компонентом клеточной стенки является пептидогликан – 40–90 %. С пептидогликаном связаны тейхоевые кислоты (имеются только у грамположительных бактерий). Тейхоевые кислоты – это полимеры из 8 – 50 остатков глицерина или рибита (пятиатомного спирта), связанные между собой фосфодиэфирными связями. Некоторые свободные гидроксильные группы в молекулах этих спиртов замещены остатками Д-аланина, глюкозы, N-ацетилглюкозамина и других сахаров. Тейхоевые кислоты пронизывают пептидогликановый слой, ковалентно связываются с пептидогликаном, достигают внешней поверхности клеточной стенки и являются антигенами, сахарные компоненты выполняют роль рецепторов для бактериофагов. Дополнительными компонентами могут быть тейхуроновые кислоты (это полимеры гексуроновых кислот) и липотейхоевые кислоты.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 |
