


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Разная степень отрицательной корреляции обнаружена между величиной удоя и способностью к откорму коров, между густотой и длиной шерсти овец, между тониной ее и длиной Величина положительной и отрицательной корреляции изменяется в зависимости от направления отбора, условий кормления и содержания животных.
Многие признаки коррелируют с другими непрямолинейно с увеличением одного из них закономерно возрастает другой лишь до определенного оптимума, после чего увеличение первого признака все в меньшей степени связано с изменением в ту же сторону второго; в какой-то момент корреляция отсутствует, а затем она может даже стать отрицательной. Такой характер корреляций наблюдается, например, между величиной удоя и живой массой коров, между продолжительностью сухостойного периода и удоем.
Вычислять коэффициенты корреляции по признакам, связь которых криволинейная, не имеет никакого смысла. Некоторые признаки, которые наследуются независимо друг от друга при разных генетических характеристиках стад и результатах отбора и подбора, ранее применявшихся, могут показывать как положительную, так и отрицательную корреляцию и ее отсутствие. Это часто наблюдается при установлении величины корреляции между удоем коров и жирностью молока.
В данном случае показатели коэффициента корреляции имеют малое практическое значение, так как по ним нельзя рассчитывать, что отбор по одному признаку повлияет на изменение другого. В каждом стаде могут быть животные, относящиеся к разным наследственным типам по сочетанию обильномолочности и жирномолочности, в том числе и такие, у которых при высоких удоях отмечается высокое содержание жира в молоке. Закрепление отбором в стаде такого типа, безусловно даст сдвиг по обоим признакам в сторону их повышения. Отсутствие корреляции между удоем и содержанием жира в молоке или отрицательная корреляция между этими признаками лишь фиксирует результаты отбора и подбора, применявшихся в прошлом.
Иногда при отборе учитывают и косвенные признаки, не имеющие прямого отношения к тем ценным свойствам, которые хотят улучшить. Например, по данным , и др., была установлена положительная корреляция между числом потовых желез на 1 мм2 кожи уха телочек и их будущей молочностью. Хотя и невысокая, но положительная корреляция установлена между молочностью коровы и длиной грудной клетки, длиной и толщиной хвоста, шириной между ребрами, углом между ребрами и позвоночником. Такая связь косвенных признаков с основными продуктивными качествами неслучайна и обусловливается общими для главных и косвенных признаков конституциональными и физиологическими особенностями.
Чем выше положительные корреляции между признаками, тем проще и успешнее их используют при отборе. Когда имеет место отрицательная корреляция между хозяйственно полезными признаками, требуется длительная племенная работа, одновременная селекция по обоим признакам с целью постепенной перестройки имеющейся отрицательной корреляции между ними. Коэффициент корреляции, указывая на степень связи между признаками, однако не дает ответа на вопрос, на сколько единиц изменится один признак при изменении другого. Более широкую информацию о связи признаков дает регрессионный анализ, при котором вычисляют специальную величину — коэффициент регрессии (R).
Повторяемость признаков. Под повторяемостью признака понимают степень соответствия между оценками по нему животного, произведенными в разное время. Например, между удоем коровы за I лактацию и последующей ее продуктивностью; между живой массой животного в раннем возрасте и массой во взрослом состоянии; между удоем коровы за первые отрезки лактации и за 305 дней лактации. Степень повторяемости признака имеет важное значение для отбора: чем она больше, тем надежнее будет отбор по первым оценкам, тем раньше можно определить племенную ценность животного. Установлено, чем в большей мере тот или иной признак зависит от условий кормления и содержания животных, тем ниже степень повторяемости. Выравненность хозяйственных условий в разный периоды оценки признака повышает степень его повторяемости.
Более высокая, или полная, повторяемость характерна для морфологических (экстерьерных) и некоторых качественных показателей, менее высокая — для количественных признаков.
Степень повторяемости признака может быть измерена коэффициентом корреляции между сопоставленными величинами. Установлена высокая повторяемость удоев коров за первые 3 месяца лактации и удоев за 305 дней (от 0,80 до 0,90), за смежные лактации (от 0,37 до 0,60), за первые 3 лактации и их пожизненной продуктивностью (от 0,82 до 0,91). Повторяемость этих показателей в условиях выровненного по годам кормления выше (от 0,60 до 0,75). Небольшая величина повторяемости обнаружена между живой массой телят при рождении и массой во взрослом состоянии (r=0,19), коэффициент корреляции между этими же показателями у ягнят 0,12—0,28.
Генетические основы отбора. Все признаки и особенности животных развиваются в определенных условиях внешней среды на основе тех возможностей, которые закладывались в половые клетки, образующие зиготу, в виде генетической информации, внесенной родительскими организмами через ДНК и РНК. Поэтому генетика как наука о наследственности живых организмов и закономерностях ее изменения занимает одно из ведущих положений в теоретических основах селекции. Особое значение в настоящее время приобретает популяционная генетика, на достижения которой опирается крупномасштабная селекция.
Понятие «популяция» было предложено в 1907 г. В. Иогансеном. Она представляет собой группу животных (или растений) одного вида, распространенную на определенной территории и способную к самостоятельной эволюции. В качестве популяции условно можно рассматривать любую породу или внутрипородный зональный тип животных, если их представителей разводят «в себе». Такие группы объединяет некоторая генетическая общность. Основой популяционной генетики служит закон Харди-Вайнберга, по которому свободно размножающаяся популяция находится в равновесии и в ней сохраняется определенная генетическая структура. Основной движущей силой изменения такой структуры является отбор. Следовательно, популяционная генетика имеет дело не с отдельными особями, а с большими группами животных, и основным методом ее исследования служит математическая статистика, опирающаяся на закономерности случайного распределения большого числа вариантов с широким применением компьютеризации, что позволяет устанавливать параметры изменения популяции в процессе смены поколений, разрабатывать на ЭВМ эффективные программы селекции.
Установлено, что наследование основных селекционных признаков имеет полигенный характер, а также значительное воздействие на них целого комплекса факторов внешней среды, что усложняет процесс наследования и создает высокую степень изменчивости. Вследствие этого морфологические, физиологические и продуктивные качества животных при сходном генном комплексе, то есть при сходных наследственных особенностях, могут быть весьма различны.
Всю совокупность внешних особенностей и продуктивных качеств животных называют фенотипом, а комплекс наследственных факторов, всю сложную структуру генетической информации, переданной через половые клетки родителей, — генотипом. Термины эти, употребляемые в племенном деле, заимствованы из генетики.
Оценку и отбор по фенотипу называют массовым отбором, а оценку и отбор по генотипу — индивидуальным. Строгих разграничений между этими формами отбора не должно быть. Генотип проявляет себя как единая, целостная система, регулирующая все процессы развития. По В. Иогансену, генотип означает норму реакции данного организма на факторы среды в течение онтогенеза. Он обусловливает не только развитие разносторонних особенностей самого животного, но и сохранение их в той или иной степени в потомстве.
Фенотип и генотип животного находятся в тесной взаимозависимости: развитие всех фенотипических признаков обусловлено определенным генотипом.
Оценка генотипа животного при отборе проводится тремя способами: по его фенотипу, по фенотипу его предков и близких родственников, по фенотипу его потомства. В племенной работе эти оценки не взаимоисключают, а дополняют друг друга. Следовательно, оценка генотипа, какими бы методами она не определялась, проводится по фенотипу. Тем не менее при сложности наследования хозяйственно полезных признаков животных фенотип не всегда и не в полной мере отражает наследственные свойства животного, его генотип. И нередко даже выдающиеся по продуктивным качествам животные дают весьма посредственное потомство.
Практика племенной работы показывает, что весьма нелегко сохранить в потомстве качества особо ценных родителей, так как влияние их как бы нейтрализуется действием наследственных факторов других менее ценных предков. Еще в конце прошлого века Ф. Гальтон, изучая статистическим методом явления наследственности, установил, что закономерным характером наследования качества родителей является тенденция возврата к средним показателям. Сущность этой закономерности состоит в том, что у лучших родителей дети оказываются несколько хуже их, а у худших — несколько лучше. Такое явление назвали законом регрессии (или закон Гальтона), который нашел подтверждение во многих более поздних исследованиях. Причем полного возврата к средним показателям стада (популяции) обычно не наблюдается, и степень регрессии бывает различной.
Изучая наследование молочной продуктивности у коров красной датской породы по материалам IV и V томов племенной книги, установил следующие закономерности. Оказалось, что дочери от лучших по продуктивности матерей имели удои ниже их, но выше по сравнению с коровами всей группы. А дочери от худших матерей, превышая их продуктивность, имели удои ниже, чем в среднем по всей группе. Аналогичное явление наблюдал , изучая наследование содержания жира в молоке коров бурой латвийской породы.
Описанные особенности наследования величины удоя и жирности молока наглядно показывают, что если оставлять для размножения (на племя) только потомков лучшей части стада, то в среднем каждое новое поколение будет лучше предыдущего по тем признакам и свойствам, по которым ведется отбор. Чем интенсивнее идет отбор в одном и том же направлении, тем в большей степени стадо или порода насыщается наследственными задатками лучших предков, и средние показатели каждого нового поколения сдвигаются в лучшую сторону, обеспечивая непрерывное совершенствование животных. Следовательно, от интенсивности отбора зависит степень возврата к среднему, и чем эта степень меньше, чем выше эффективность селекции.
Несмотря на закономерное действие регрессии, отбор лучших по селекционным признакам животных не остается безрезультатным, обеспечивая сдвиг средних показателей стада со сменой поколений, и тем в большей мере, чем интенсивнее идет отбор. Схематично это показано на рис. 35.
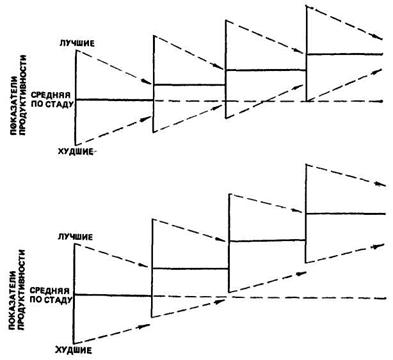
Рис. 35. Сдвиг средних показателей продуктивности стада со сменой поколения, отбором лучших животных при разной степени регрессии
При использовании массового отбора прогресс любого стада зависит от того, какая часть или в какой степени высокие продуктивные качества родителей будут унаследованы потомством. Поэтому практически важно установить степень надежности того, что отбором лучших животных по фенотипу будут «улавливаться» и лучшие генотипы.
Выявление доли влияния генотипа родителей на количественный признак потомства было начато работами С Райта и затем продолжено работами Р. Фишера, Д. Лаша, И. Лернера, Д. Фальконера. В основу анализа фенотипической изменчивости признака были положены разработанные С Райтом показатели корреляции между фенотипом и генотипом родителей и потомства. Замкнутая цепь этих связей видна на рис. 36. При этом допущены теоретические предположения, что популяция находится в генетическом равновесии, количественные признаки определяются аддитивными (суммирующего действия) генами и что корреляция между генотипом родителя и генотипом потомства равна 0,5. Величина h — это коэффициент пути от генотипа к фенотипу. По схеме связи между генотипом и фенотипом родителей и потомка (см. рис. 36) можно получить следующее выражение корреляции между фенотипом родителей и фенотипом потомка r=hr, между генотипом родителей и генотипом потомка r=0,5h2, после преобразования равенства h2=2r между фенотипом родителей и фенотипом потомка. Величину h2 С. Райт назвал коэффициентом детерминации фенотипа генотипом.
Американским ученым Д. Лашем (1939) введено понятие «наследуемость признаков» и величина h2 названа коэффициентом наследуемости. Используется он для определения в общей фенотипической
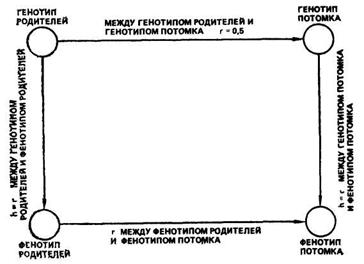
Рис. 36 Замкнутая цепь коррелятивных связей между генотипами и фенотипами родителей и потомка (по Райту)
изменчивости той доли, которая обусловливается наследственными различиями организма (его генотипом). Существуют разные способы вычисления коэффициента наследуемости:
1) h2=2r между показателями одного и того же признака родителей и потомков. Если продуктивность получают от животного только одного пола, например молочная продуктивность коров, то коэффициент наследуемости выражается удвоением коэффициента корреляции между продуктивностью матерей и дочерей (h2=2rМд)
С А Рузский (1977), считая, что удвоение коэффициента корреляции обычно приводит к завышению h2, а иногда и к явно ошибочным результатам, когда h2 выражается величиной, превышающей единицу, предложил за коэффициент наследуемости брать коэффициент корреляции между родителями и потомством без его удвоения, то есть h2=2rмд;
2)h2=2R между показателями одного и того же признака родителей и потомства. Эта формула разработана Д. Лашем. По ней коэффициент наследуемости равняется удвоенному коэффициенту регрессии между показателями признака родителей и потомства;
![]()
где коэффициент наследуемости h2 равняется отношению показателя дисперсии, вызываемой генетическими факторами (Сх), к общей фенотипической дисперсии признака (Су);
![]()
где Ммл и Ммх — средние показатели лучших и худших матерей по сравнению со средним по стаду, Мдл и Мдх — средние показатели того же признака у дочерей, полученных от лучших (л) и худших (х) матерей.
Предлагались и другие методы вычисления коэффициента наследуемости (по корреляции между сибсами и полусибсами, методом дисперсионного анализа и др.).
Величину коэффициента наследуемости выражают в долях единицы или в процентах. Например, если по постоянству лактационной кривой у коров h2 = 0,12, или 12 %, то это означает, что характер лактационной кривой у коров-матерей на 12 % обусловлен наследственностью и в такой же мере унаследован их дочерьми. Чем выше коэффициент наследуемости тех или иных признаков, тем в большей степени изменчивость их определяется наследственными различиями и тем более эффективным будет массовый отбор по этим признакам
В нашей стране и за рубежом широко ведутся генетикостатистические исследования популяций и накоплено уже большое количество данных о степени наследуемости самых различных селекционных признаков животных. Но при этом отмечены весьмa большие различия в показателях коэффициента наследуемости даже одних и тех же признаков.
Признак | коэффнциент наследуемости, % |
Молочный и молочно-мясной крупный рогатый скот | |
Величина удоя | 4—60 |
Содержание в молоке: жира | 17—70 |
белка | 45—70 |
Характер лактационной кривой | 10—30 |
Скорость молоковыведения | 15—45 |
Мясной крупный рогатый скот | |
Живая масса, при рождении | 11—53 |
в 12—15 месяцев | 36—94 |
Убойный выход | 25—73 |
Площадь «мышечного глазка» | 50—73 |
Плодовитость коров | 10-22 |
Свиньи | |
Живая масса | 0—56 |
Длина туловища | 40—86 |
Число поросят при рождении | 6—34 |
» сосков | 10—60 |
Среднесуточный прирост | 14—77 |
Скороспелость | 4—39 |
Молочность | 12—61 |
Толщина шпика | 25—84 |
Куры | |
Яйценоскость за первый год | 11—47 |
Масса яиц | 33—80 |
Выводимость | 3—20 |
Жизнеспособность молодняка и взрослой птицы | 3—15 |
Живая масса взрослых кур | 50—65 |
2—3-месячных цыплят | 25—50 |
Оперяемость цыплят в 2-месячном возрасте | 25—40 |
Установлено, что степень наследуемости будет тем выше, чем однороднее и стабильнее условия существования животных разных поколений. Обычно чем выше уровень кормления животных и их продуктивность, тем больше коэффициент наследуемости. Ф Ф. Эйснер писал: «Наследственность всегда проявляется в конкретных условиях. Вне среды — наследственность лишь абстрактное понятие. Генотип определяет норму реакции организма на внешние условия. Меняются условия — неизбежно меняется и норма реакции. Значит, отделять влияние наследственности от влияния среды можно лишь с большой долей условности и лишь в ограниченных пределах колебания внешних факторов». Иллюстрируется это примером вычисления коэффициента наследуемости удоя за I лактацию у симментальского скотам в 4 племзаводах Украины при разных условиях кормления матерей и дочерей Когда и матери, и дочери находились в плохих условиях кормления, коэффициент наследуемости колебался в пределах 0,024— 0,238; когда матери были в худших, а дочери в лучших условиях, величина коэффициента наследуемости была 0,160—0,274; но когда и матери, и дочери находились В лучших условиях, величина коэффициента наследуемости составляла 0,394—0,584.
Величина коэффициента наследуемости значительно варьирует в зависимости от породы (популяции), местных особенностей породных групп, генеалогической структуры стада, уровня и направления племенного отбора, применявшихся методом разведения, систем спаривания и других особенностей. В табл. 25 приведены данные о различии показателей коэффициента наследуемости удоя и содержания жира в молоке у инбредных и аутбредных коров швицкой породы.
25. Коэффициент наследуемости продуктивности коров-первотелок (по данным Солдатова)
Показатели | Число коров | Коэффициент наследуемости, % |
Удой за лактацию: аутйредные | 173 | 6,78 |
инбредные | 49 | 23,48 |
Содержание жира в молоке: аутбредные | 178 | 18,42 |
инбредные | 49 | 47,84 |
Примером больших различий величин коэффициента наследуемости одних и тех же признаков в различных стадах могут служить данные, приведенные в табл. 26,
В связи с различием методов вычисления коэффициента наследуемости и влияния на его величину большого числа факторов абсолютные величины h2 имеют несколько условный характер. Но для разработки программ селекционного улучшения стада важно даже примерное значение коэффициента наследуемости, позволяющее устанавливать различие в степени наследуемости отдельных признаков и использование этого коэффициента как ориентира при перспективном планировании племенной работы.
Несмотря на большое варьирование величины коэффициента наследуемости одного и того же признака, выявлена некоторая закономерность, заключающаяся в том, что хозяйственно полезные признаки животных различаются по степени наследуемости. Так, у крупного рогатого скота степень наследуемости скороспелости и мясности выше, чем молочности и состава молока.
26. Коэффициент наследуемости продуктивности коров черно-пестрой породы в среднем за три лактации (по данным Эрнста, Чемма)
Хозяйство | Число пар Мать-дочь | Коэффициент наследуемости, % | |
удой за лактацию | содержание жира молоке | ||
«Молочное» (Вологодская область) | 190 | Близко к 0 | Близко к 0 |
«Холмогорка» (Московская область) | 410 | 9,0 | 36,0 |
«Исток» (Свердловская область) | 328 | 16,8 | 16,8 |
«Петровское» (Московская область) | 133 | 22,6 | 42,4 |
«Константинове» | 209 | 24,0 | 63,0 |
(Московская область) «Никоновское» (Московская область) | 338 | 27,2 | 16,8 |
Совхоз имени В И Ленина | 144 | 29,6 | Близко к 0 |
(Калининская область) «Горки П» (Московская область) | 208 | 34,6 | 50,4 |
Учхоз КСХИ (Кировская область) | 121 | 36,0 | Близко к 0 |
«Первомайское» (Московская область) | 284 | 48,6 | 21,4 |
Наследуемость содержания жира и белка в молоке выше, чем наследуемость величины удоя. По данным , наследуемость живой массы и настрига шерсти у помесных кросс-бредных овцематок с тониной шерсти 50-го качества (h2=19,2 и 22,8%) оказалась более высокой, чем у овец с другими качествами шерсти. У овец с шерстью 48—46-го качества наследуемость длины и тонины ее (h2=25,6 и 23,6%) была выше, чем у овец с другими качествами шерсти, и выше, чем наследуемость в этой же группе живой массы и настрига шерсти (h2=15,1 и 16,6%) Наследуемость молочности и крупноплодности у свиноматок выше, чем плодовитости. У кур степень наследуемости толщины скорлупы яиц меньше, чем формы и качественных показателей яиц.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 |
