


Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
В работе , проведенной на большом поголовье лошадей (402 головы) орловской породы Московского конзавода № 1 и Хреновского завода № 10, были изучены различные типы трансферринов и обнаружено 6 аллелей.
Установлена достоверная разница в резвости и промерах лошадей с различными генотипами по трансферрину. Было также выяснено, что различные сочетания родительских пар по типу трансферрина влияют на качество потомства, их резвость и развитие. Лучшая резвость была при сочетании
♂HOX♀OF — резвость потомства 2 мин 13,0 с, ♂DHX♀DO — 2 мин 17,3 с, худшая резвость при сочетании ♂DHX♀DO— 2 мин 24,9 с и ♂FOX♀FH — 2 мин 24,8 с.
Установлена связь особенностей групп крови с энергией роста и величиной живой массы в различные возрастные периоды у, мясных пород крупного рогатого скота, а также со средним суточным приростом и оплатой корма у свиней. Американский ученый Станевильд с сотрудниками выявил следующую закономерность: овцы, гетерозиготные по некоторым локусам групп крови, отличались большой живой массой и лучше выраженными мясными признаками. Это же подтвердилось исследованиями и на других видах животных. Гетерозиготные коровы по антигенным факторам оказались более продуктивными по сравнению с их гомозиготными сверстницами.
отмечает, что у свиней, гетерозиготных по некоторым системам групп крови, плодовитость выше, чем у гомозиготных. Гетерозиготные животные на откорме по сравнению с гомозиготными отличались лучшей скороспелостью, более высокими мясными качествами и лучшей оплатой корма продукцией. Опыты, проведенные в Румынии на трех породах овец, показали, что особи, гетерозиготные по типам гемоглобина, характеризовались большей шерстной продуктивностью, чем овцы гомозиготные.
При изучении полиморфных систем белков молока (Меркурьева, Скрипниченко) выявлено заметное породное различие в концентрации генов и генотипов по четырем изученным локусам. При исследовании аллелей в локусах белков молока установлено, что у четырех пород крупного рогатого скота (черно-пестрая, холмогорская, голландская и айрширокая) обнаружены одинаковые аллели (А и В) по локусу β—Lg (бета-лактоглобулину), по β- и К — Сп (бета - и каппа-казеину), а по локусу αS1—Сn (альфа-эс-один-казеину) аллели В и С найдены у черно-пестрого, холмогорского и голландского скота. У айрширов в этом локусе обнаружен мономорфизм, отсутствовали аллели А, С, D, что является породной особенностью популяции. Установлены большие породные различия в частоте аллелей. Так, по β—Lg у коров черно-пестрой породы высокая частота оказалась по аллелю A, а у айрширского, холмогорского и голландского скота — по аллелю В.
По каждой из пород и по каждому локусу был проведен по-пуляционный анализ на выявление генетической структуры популяции по полиморфным системам белков молока. С этой целью определяли встречаемость и соотношение генотипов. Оказалось, что процент гетерозиготных генотипов был во всех четырех стадах более низким, чем процент гомозиготных генотипов.
Определенная связь генотипа группы крови выявлена по поражаемости коров айрширской породы маститом. Наибольший процент заболеваний (до 23—33%) маститом выявлен у коров — носительниц генотипов по локусам трансферрина AD, церуло-плазмима АА, амилазы ВС, каппа-казеина АА. Установлена наследственная обусловленность заболевания маститом в дочернем потомстве 12 быков айрширокой породы. Изучение генетических основ резистентности молочного скота к маститам, кур к болезни Марека и пуллорозу, свиней к инфекционному атрофическому риниту принимает сейчас широкий размах.
Большой научно-практический интерес к проблеме полиморфизма биологических структур сельскохозяйственных животных привел к тому, что создано Международное общество по изучению групп крови животных, на которое возложен ряд важных функций по сравнительной оценке качества реагентов отдельных видов животных соответствующей международной классификации, комплектование банка моноспецифических сывороток крови и вариантных типов белков крови и молока, координация исследований по иммуногенетическим и другим вопросам. Накоплено значительное число данных, показывающих, что гетерогенный подбор по определенным генетическим системам антигенных факторов ведет, к гетерозису. Фирма «Тонбер» в Англии и фирма «Хай-Лайн» в США практикуют маркировку линии по антигенным факторам. В Англии в результате селекции по иммунобиологическим показателям в течение пяти лет яйценоскость кур повысилась на 28 яиц в год, затраты корма уменьшились на 16%, а падеж снизился на 30%.
Ферменты крови. По ним можно судить о продуктивных и племенных качествах животных в раннем возрасте. Ферменты являются биологическими катализаторами, участвующими во всех жизненно важных процессах, протекающих в организме. Действие генов осуществляется через ферменты по такой схеме: ген — фермент — биохимическая реакция — признак. Известно, что на формирование сложных хозяйственно полезных признаков оказывают влияние одновременно много генов. В связи с этим дает такую схему взаимосвязи наследственных задатков, ферментов и признаков: много генов — много ферментов — один признак. Биохимической генетикой установлено, что уровень активности ферментов крови обусловлен наследственными факторами. В настоящее время характер наследования 16 типов ферментов изучен (Смирнов) у человека, птицы, кроликов, лошадей, свиней. В эти типы входят следующие ферменты: щелочная и кислая фосфатаза, амилаза, аминотрансфераза, эсте-раза, каталаза и др. Они служат своеобразными биохимическими тестами, по которым можно в молодом возрасте определить будущую ценность животных. Поэтому изучение генетических систем, контролирующих ферменты крови, и систем, определяющих особенности индивидуального развития животных (скорость роста, оплата корма, продуктивные качества), имеет большое значение.
Активность ферментов крови у крупного рогатого скота в зависимости от пароды и возраста животных изучали курьева и (табл. 7).
7. Различия в активности ферментов крови первотелок разных пород
(удои 44кг за лактацию, жирность молока 4,28 %)
Порода | Амилаза, % | Щеточная фосфатаза, мг% | Альдолаза. ед | Аспартатами- нотрансферазa, ед | Аланинамино- трансфераза, ед. |
Айрширская Голландская | 12,86 13,75 | 1,33 2,17 | 23,62 24,82 | 47,82 47,40 | 21,10 19,22 |
В ходе лактации активность ферментов изменяется. Наиболее высокие уровни по альдолазе отмечены в крови коров на втором месяце лактации. Активность аспартат - и аланинаминотрансферазы повышается со второго до четвертого месяца, а затем снижается. Связь среднелактационного показателя активности ферментов с удоем, выраженная коэффициентом корреляции, колебалась от 0,3 до 0,8. Корреляция с содержанием жира и белка была положительной, а с удоем — отрицательной. Установлена положительная корреляция между аланином (АЛТ) и процентом жира (r=0,629) и белка (r = 0,678) в молоке айрширских коров; между удоем и этим ферментом корреляции не обнаружено. Активность ферментов сыворотки (аминотрансфераза) крови оказывает существенное влияние на плодовитость кроликов, свиней, овец. приводит интересные данные о различной эффективности подбора пар у кроликов. На рис. 26 видно, что в одинаковых условиях кормления и содержания инбредный самец № 000 с высокой активностью аминотрансфераз (ACT и АЛТ) в сыворотке крови был спарен с тремя самками (№ 000, 1854 и 358), у которых отмечена высокая, средняя и низкая активность этих ферментов. В первом случае получено 8 крольчат, во втором — 6, в третьем — 3. Сохранность крольчат была лучшей в группе при гомогенном подборе родителей по высокой ферментативной активности. Установлена связь активности аминотрансфераз с энергией роста и живой массой свиней, овец.
В качестве интерьерных показателей в последние годы служат данные, характеризующие особенности хромосомного аппарата соматических клеток. Новым направлением цитогенетики сельскохозяйственных животных является изучение хромосомного полиморфизма породных и индивидуальных различий размеров и внутренней структуры хромосом. Обнаружены такие перестройки хромосом у крупного рогатого скота, овец и других видов животных, как транслокации, инверсии, чепы (щели), хрома-тидные разрывы и др., которые вызывают патологический обмен веществ, уродство, болезни, раннюю эмбриональную смертность, бесплодие.
Одной из наиболее распространенных форм кариотипической изменчивости хромосом является транслокация (центрическое слияние хромосом), описанная Робертсоном. Данная хромосомная мутация обнаружена у 40 пород крупного (рогатого скота и других видов животных.
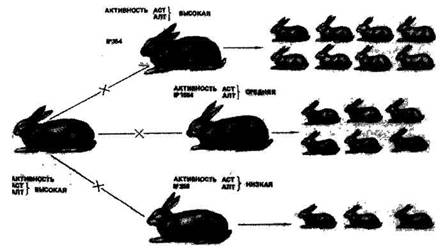
Рис 26. Влияние подбора кроликов по активности ферментов сыворотки крови на их плодовитость (по Смирнову)
Носителями таких хромосомных аномалий являются производители. Новейшими исследованиями , , установлено отрицательное влияние робертсоновских транслокаций на продуктивность и плодовитость коров, унаследовавших от отца эту аномалию кариотипа. Прослеживается также связь этих транслокаций с предрасположенностью животных к лейкозу и другим заболеваниям.
Чтобы очистить племенные стада от генетического брака, необходимо как обязательное мероприятие ввести кариологнчеокое обследование всех быков-производителей (на станциях искусственного осеменения) и ремонтных бычков (выращиваемых для племенных целей). В 1976—1987 гг, были проведены в MBA им. (кафедра генетики и разведения с.-х. животных) исследования 6 плановых пород крупного рогатого скота, зебу, буйволов и гибридов между ними, а также пород свиней и дикого кабана (ЧСФР).
Цитогенетические исследования представляют интерес в связи с изучением видовой принадлежности животных и выявлением особенностей у гибридов. Так, при изучении хромосом зебу и их гибридов с крупным рогатым скотом обнаружены различия в морфологии игрек-хромосомы, установлена более высокая митотическая активность у гибридных животных по сравнению с показателями исходных форм, что может указывать на проявление гетерозиса. Изучение кариотипов буйвола кавказской породы и гибридов (кавказский буйволXиндийский буйвол породы мурра) дало возможность впервые установить их нормальный кариотип. Исследование морфологических структур хромосом этого скота выявило связи между кариотипом и резистентностью, а также адаптивными возможностями животных, что имеет немаловажное значение в селекционной работе.
Контрольные вопросы. 1. Что вы знаете о типах конституции животных? 2. Расскажите о классификации типов конституции сельскохозяйственных животных. 3. Охарактеризуйте роль наследственности и условий среды в формировании конституциональных типов. 4. Дайте характеристику крепкого, старого (рыхлого) и сухого типов конституции. 5. Расскажите о связи конституции с различными проявлениями жизнедеятельности организма (здоровье, продуктивность). 6. Что такое кондиция? 7. Охарактеризуйте методы оценки экстерьера сельскохозяйственных животных. 8. Какие зоотехнические задачи решаются с помощью экстерьерной оценки животных? 9. Что такое интерьер животных? 10. Охарактеризуйте методы интерьерной оценки.
Глава IV. РОСТ И РАЗВИТИЕ ЖИВОТНЫХ
УЧЕНИЕ ОБ ОНТОГЕНЕЗЕ
Изучению индивидуального развития животных (онтогенеза) были посвящены исследования многих деятелей науки. Особенно интенсивно и плодотворно теория онтогенеза стала разрабатываться в последние 15—20 лет в связи с развитием молекулярной биологии и генетики. Молекулярно-генетичеокие методы исследования углубили познания закономерностей роста и развития животных, создали новые возможности для разработки методов управления этими процессами.
Знание индивидуального развития организма необходимо прежде всего потому, что в процессе роста и развития животное приобретает не только породные и видовые признаки, но и присущие только ему особенности конституции, экстерьера, продуктивности. В онтогенезе осуществляется наследственная преемственность и изменчивость признаков родителей, он протекает в результате действия внутренних природных факторов организма и условий внешней среды. Становление всех хозяйственно полезных признаков животных, таких как молочность, яйценоскость, настриг шерсти, скорость бега, происходит благодаря развитию наследственной основы организма в конкретных условиях среды. Иными словами, чтобы получить высокопродуктивное животное нужно уметь его выращивать. Онтогенез состоит из двух основных процессов: роста и развития. Хотя эти понятия и взаимосвязаны, но не тождественны
Под ростом понимают процесс увеличения размеров организма, его массы, происходящий за счет накопления в нем активных главным образом белковых, веществ.
Рост сопровождается не только увеличением массы, но изменением пропорций тела, обусловливающим новые качества. В основе роста животных лежат при различных процесса, деление клеток, увеличение их массы и объема, увеличение межклеточных образований
Понятие роста организма по разному толковалось многими учеными В. Ру, Г. Дриш, К. Сент-Илер под ростом понимали всякое увеличение размеров тела, его объема, массы.
— один из крупнейших эволюционистов-биологов, истолковывал рост как увеличение активных частей организма, при котором количество свободной энергии в организме возрастает, но не всякое увеличение объема можно назвать ростом. на основе глубоких исследований онтогенеза ростом называл увеличение массы клеток организма, его тканей и органов, линейных и объемных их размеров, осуществляемых за счет количественных изменений в результате стойких новообразований живого вещества.
Увеличение массы в результате накопления резервных жировых веществ или воды в теле животного не характеризует процесс роста. Приведем два примера. При откорме закончившая рост свиноматка жиреет, ее живая масса увеличивается. Взрослые курдючные овцы на обильных пастбищах накапливают большие запасы жира в курдюке (до 35—40 кг). В данных случаях увеличение массы организма не является собственно процессом роста. Рост молодых животных происходит вследствие активного обмена веществ, накопления в организме белка; у взрослых животных этот процесс имеет иной характер (происходит жирообразование). Рост тела животного непосредственно зависит от преобладания процессов синтеза, ассимиляции над процессами диссимиляции (распада) веществ.
В науке пока нет единой точки зрения на взаимосвязь между ростом и развитием. Например, крупнейший английский биолог Д. Хеммонд считает развитие производным процесса роста. Ряд ученых под развитием понимают только усложнение структуры организма, дифференциацию органов и тканей, отводя росту главную роль в развитии. справедливо подчеркивает, что рост животного — один из основных показателей его индивидуального развития. указывал, что развитие организма осуществляется в результате действия двух основных процессов: роста и дифференциации. Направление и формы последней контролируются, с одной стороны, всем последовательным ходом онтогенеза, определенным генотипом и являются результатом возникших с самого начала развития (дробление яйца) связей между частями развивающегося зародыша; с другой стороны, мощное формирующее влияние на конкретный ход развития оказывают условия среды, окружающей развивающийся организм.
Современные генетические и эмбриологические исследования внесли много нового в теорию онтогенеза, в процессы управления индивидуальным развитием, вычленили состояние клеток на ранних стадиях развития и их подавления, выяснили значение мигрирующих генетических элементов в онтогенезе, определили роль генов-селекторов в процессе дифференциации, репрессии и депрессии как средств регуляции и развития.
ОСОБЕННОСТИ РОСТА И РАЗВИТИЯ ЖИВОТНЫХ
Взаимосвязь между процессами роста и развития — это взаимосвязь между количественными и качественными изменениями, происходящими в организме в процессе онтогенеза. Нельзя говорить об изолированности процессов роста и развития животных, оба они взаимосвязаны и взаимообусловлены.
Под развитием животного понимают процесс усложнения структуры организма, специализацию и дифференциацию его органов и тканей. Иными словами, развитие — это качественные изменения содержимого клеток, органообразовательные процессы, которые проходит каждый организм от оплодотворенного яйца до взрослого, способного к размножению и сходного в основных чертах с родительским организма.
Для развития животного характерны следующие важнейшие особенности:
1) специализация клеток, органов и тканей в выполнении определенной функции в организме;
2) возникновение новых и усложнение функций органов и тканей (морфогенез). Дифференциация и специализация органов и
тканей не сопровождается независимой жизнедеятельностью частей организма, наоборот, это приводит к объединению и соподчинению деятельности органов и тканей. В последние годы процесс
клеточной дифференциации объясняется с позиций теории дифференцированной активности генов, которая является одной из
наиболее важных обобщающих теорий в биологической науке
XX в. Согласно этой теории специализация клеток — результат
действия соответствующих групп генов, характерных для каждого типа клеток;
3) объединение и взаимосвязь развития различных органов и
тканей. У млекопитающих и птиц эту функцию (интеграции) выполняют нервная и эндокринная системы, ферменты и кровь;
4) приспособление организма к конкретным условиям внешней среды;
5) периодизация индивидуального развития животного.
В процессе онтогенеза животных в одних случаях усиленная дифференциация сопровождается снижением скорости роста, в других случаях бурный рост связан с замедленным развитием организма. Можно одновременно наблюдать интенсивность роста и развития или обоюдную депрессию этих процессов при неблагоприятных условиях среды. В отличие от млекопитающих у насекомых отмечается резкая периодизация онтогенеза, взаимоисключающая процессы развития и роста. Интенсивный рост у насекомых идет только между линьками, а развитие — во время линьки. В организме животных эти процессы протекают параллельно, обусловливая друг друга.
Развитие организма начинается с оплодотворения яйцеклетки и формирования зиготы (оплодотворенная яйцеклетка), которая представляет собой сложное неоднородное биологическое образование Зигота включает в себя хромосомные и нехромосомные системы отца и матери.
Ряд ученых на основании современных цитогенетических исследований считают, что потенции к развитию зиготы в известной степени зависят от градиентов эндоплазматических материалов и неоднородности эктоплазмы самой зиготы. Это явление определяет как градиентное отношение в онтогении. Клетки, получающие во время дробления зиготы нехромосомный материал, формируют впоследствии различные ткани зародыша. Расщепление нехромосомного материала во время дробления зиготы — первая стадия дифференциации, и только позднее причиной ее становятся изменения ядерного материала. Гены активно включаются в процесс развития, начиная с фазы гаструляции, создающей химическую дифференциацию зародыша. В дальнейшем любые изменения, происходящие в формирующемся организме, обусловливаются генетическими факторами
Исследования показали, что существует не только прямая, но и обратная связь между развивающимся организмом и хромосомным аппаратом, т. е. активность генов хромосом зависит от их окружения (состояния нуклеоплазмы и цитоплазмы с их макромолекулами). Накоплены убедительные экспериментальные данные, указывающие на то, что преемственность между поколениями обеспечивается не только ядерным материалом, но и цитоплазматическими элементами клетки (ядерная оболочка, мембрана, митохондрия). Обнаружены и своеобразные рекомбинации цитоплазматичеоких генов. Цитоплазма является активным участником в передаче ядерной генетической информации в ряде поколений. Наблюдается и неядерный тип наследования соответствующих признаков. Каждый организм наследует от своих близких и далеких предков способность определенным образом развиваться и реагировать на воздействие внешней среды, формируя свои индивидуальные качества, в том числе и особенности, обусловливающие преемственность между поколениями животных данного вида и породы.
Зигота (наследственная основа) несет в себе отпечаток всей предшествующей истории развития данного вида животных, то есть его филогенеза. Напомним, что под генотипом следует понимать весь комплекс наследственной информации, определяющей генеральную линию развития организма. Именно этим и объясняется постоянство видовых, породных, линейных свойств и признаков животных. В процессе онтогенеза животного происходит как бы раскрытие его генотипа, завершающееся формированием фенотипа взрослой особи. Фенотип — это комплекс всех признаков и состояние особи в данный момент, на определенном этапе онтогенеза. Обусловлен фенотип наследственной природой организма и условиями среды.
Важнейшей биологической особенностью зиготы является способность повторять филогенез — путь исторического развития, пройденный предками. В связи с этим онтогенез (индивидуальное развитие организма) представляет собой краткое повторение исторического развития вида. Проблема соотношения онтогенеза и филогенеза была рассмотрена Ч. Дарвиным. Он считал, что эволюционные изменения видов в живой природе имеют в своей основе изменения, произошедшие в индивидуальном развитии. В 1866 г. Э. Геккель сформулировал биологический закон: онтогенез есть краткое повторение филогенеза. Он утверждал, что филогенез — единственная причина онтогенетического развития. Признание одностороннего влияния филогенеза на онтогенез ошибочная точка зрения. Крупнейший эволюционист неоднократно подчеркивал, что под филогенезом следует понимать ряд исторически отобранных онтогенезов. Онтогенез не только результат филогенеза, но и его основа. В филогенезе отбираются, концентрируются и закрепляются те изменения, которые возникали в онтогенезе многих поколений и обеспечивали возможность существования вида, его развитие. Филогенез peaлизуется в онтогенезе через наследственность и составляет основу онтогенеза, он направляет онтогенез по пути, пройденному предками.
В ходе эволюции первичная связь между поколениями, осуществляемая через половые клетки, начинает все больше осложняться различными вторичными формами связи: а) обеспечение яйцеклетки питательным материалом; б) условия внутриутробного развития; в) питание молоком матери; г) различные формы заботы о потомстве; д) передача навыков поведения путем подражания и др. Все эти вторичные формы связи советский ученый называл биологичеоким контактом поколений. Изучение данной проблемы представляет большой интерес для современной науки, в том числе и зоотехнии.
В фенотипе животного никогда не реализуются полностью все генотипические возможности организма. Полагают, что у высших животных в геноме содержится около 300 тыс. генов. Однако в каждый период онтогенеза генетическая информация считывается только с 5% генов, именно тех, которые обусловливают контроль необходимых в данный момент белков. Для реализации их нужны соответствующие условия внешней среды. Следовательно, развитие животного, формирование его признаков и свойств определяются как генотипом данной особи, так и внешними условиями, в которых проходит онтогенез.
Для онтогенеза всех видов сельскохозяйственных животных характерен ряд общих генетических, биохимических, морфологических и физиологических закономерностей. К генетическим закономерностям относятся следующие:
1) генетическая обусловленность онтогенеза, постоянство
признаков и свойств животного организма;
2) зависимость формирования фенотипа животного от его генотипа;
3) наследственная обусловленность скорости и продолжительности роста, достижение в оптимальных условиях среды определенных размеров тела животного и продолжительности его
жизни;
4) наследственная основа организма может измениться за
счет мутаций.
Биохимические закономерности онтогенеза заключаются:
1) в направленности всех биохимических процессов на повышение взаимодействия клеток, органов и тканей организма, на
создание единой саморегулируемой системы;
2) в угасании с возрастом ассимиляторных процессов, уменьшении в органах и тканях содержания воды и увеличении количества минеральных веществ;
3) в уменьшении с ходом онтогенеза отложения в организме
азотистых веществ, в результате чего происходит понижение физико-химической активности белков тела и крови, возрастает количество холестерина.
К морфологическим закономерностям относятся:
1) снижение интенсивности роста животных с возрастом;
2) зависимость темпов роста организма, отдельных органов и
тканей от условий кормления и содержания животных (закон
недоразвитости).
Физиологические закономерности онтогенеза можно заключить в следующие положения:
1) стадийный характер роста и развития (эмбриональный и
постэмбриональный периоды);
2) на каждой стадии онтогенеза особи поддерживается со
стояние подвижного равновесия ее систем и функций;
3) в процессе старения проявляются изменения в поведении
организма, снижении его жизнеспособности и приспособленности
к условиям среды.
Весь путь от оплодотворенной яйцеклетки до взрослого животного, способного к размножению и продуцированию, состоит из двух периодов: эмбрионального и постэмбрионального. Многие ученые называют их стадиями развития. Эмбриональный период животного организма включает в себя зародышевую, предплодную и плодную фазы. Зародышевая фаза продолжается от оплодотворения яйцеклетки до полной имплантации эмбриона.
В это время идет формирование и функционирование плаценты, образование систем, органов и тканей, определяющих специфические признаки данного вида и породы, особенности телосложения и жизнедеятельности животного. Предплодная фаза — от конца зародышевой до начала плодной, а плодная — от конца предплодной до рождения животного.
В зависимости от условий эмбрионального развития продолжительность зародышевой и плодной фаз может изменяться. Особенности прохождения эмбрионального развития животного обусловлены сложной взаимосвязью плода и материнского организма. Эта связь осуществляется через плаценту, которая представляет собой орган, обеспечивающий обмен веществ между матерью и развивающимся в ее теле эмбрионом, а также дыхание, питание плода, выделение продуктов обмена. Размеры плаценты и площадь котиледонного аппарата определяют в итоге размеры и жизнедеятельность новорожденных животных.
На продолжительность внутриутробного развития животных оказывают влияние наследственные факторы, а также условия кормления, содержания и использования. По данным , кобылица с удлиненным сроком жеребости (на 7—10 дней) дает более ценный в племенном отношении приплод, чем матки, у которых период жеребости укорочен Более длительное эмбриональное развитие у ряда пород и линий является наследственно обусловленным ценным селекционным признаком.
В постэмбриональный период можно выделить следующие фазы: новорожденности, молочного питания, наступления половой зрелости, функциональной зрелости (фаза расцвета всех физиологических функций и продуктивности организма) и старения, когда в организме процессы диссимиляции преобладают над процессами ассимиляции. Каждая фаза постэмбрионального развития характеризуется своими морфологическими, физиологическими и биологическими особенностями. Индивидуальное развитие животных можно представить следующей схемой (с. 104).
ИЗУЧЕНИЕ РОСТА И РАЗВИТИЯ ЖИВОТНЫХ
Определение скорости роста. Скорость роста животных в разные периоды их жизни неодинакова. Рост определяют по живой массе и промерам. Различают абсолютный и относительный прирост живой массы. Под абсолютным приростом понимают увеличение живой массы и промеров молодняка за определенный отрезок времени (сутки, декада, месяц, год), выраженное в килограммах. Абсолютный прирост животных представляет собой разницу между массой тела конечной и начальной, разделенную на число дней.


Пример. Теленок бестужевской породы при рождении весил 35 кг, а в возрасте 30 дней — 59 кг. Абсолютный суточный прирост живой массы вычисляют по формуле
A=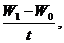
где A — абсолютный прирост, W1 — живая масса конечная; W0--живая масса начальная; t — время. В нашем примере
A=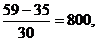
Абсолютный прирост единицы массы тела в единицу времени не может характеризовать истинную скорость роста. Для этой цели вычисляют относительный прирост, который выражают в процентах. Вычисляют его по формуле
B=
В нашем примере
B=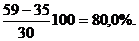
Метод вычисления относительного прироста, предложенный А. Майонотом, был усовершенствован С. Броди. При вычислении скорости роста он отнес величину абсолютного прироста (A) не к первоначальной массе (Wo), а к промежуточной величине между первоначальной и конечной Формула имеет следующий вид:
B=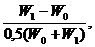
Скорость роста определяют также по формуле, предложенной С. Броди и .
Cw=
где Сw, — скорость роста; W1 — масса начальная; W2 — масса конечная; t — время; 0,4343 — основание натуральных логарифмов.
По этой формуле можно определить константу роста К= = Cw·t. Произведение скорости роста на возраст есть величина постоянная. Иначе говоря, скорость роста снижается пропорционально возрасту. Постоянство константы роста характерно только для определенных периодов жизни. Зная величину константы роста и возрастные периоды, на которые она распространяется,
8. Скорость роста животных в эмбриональный и постэмбриональный периоды (по данным Рубнера)
Вид животного | Масса | Продолжительность беременности. Мес. | Число удвоенной массы | ||||
Зиготы Мг. | при рождении животного. Кг. | Во взроcлом состоянии, кг | в эмбриональный период | в пост эмбриональный период | за весь период | ||
Лошадь | 0,60002 | 50,0 | 500,0 | 11,3 | 26,3 | 3,43 | 29,7 |
Собака | 0,4015 | 0,22 | 11,0 | 2,10 | 19,09 | 5,29 | 24,3 |
Кошка | 0,6301 | 0,087 | 3,80 | 1,87 | 17,07 | 5,79 | 22,8 |
Морская свинка- | 0,499 | 0,050 | 0,10 | 1,97 | 16,61 | 3,58 | 20,19 |
можно прогнозировать конечные показатели роста животных по отдельным отрезкам времени.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 |
