

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral

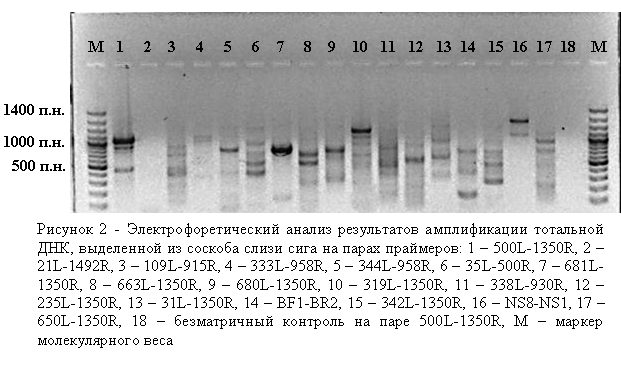
Нами ранее показано, что в результате ПЦР на консервативных праймерах получено два продукта длиной 900 и 1050 пар нуклеотидов (п. н.) (Белькова и др., 2008). Более длинный ПЦР-продукт соответствует фрагменту гена 18S рРНК исследуемого вида рыб. К сожалению, загрязнение ДНК организма-хозяина при проведении видо-специфичной амплификации на бактериальных праймерах приводит к ложно отрицательному результату. Нами использованы праймеры с разным уровнем специфичности для дальнейшей разработки методики (табл. 4). В серии экспериментов показано, что с разных матричных ДНК получается разный спектр целевых ампликонов, неспецифичные полосы могут синтезироваться с ДНК организма-хозяина (рис. 2). На следующем этапе была предпринята попытка селективной амплификации бактериальной ДНК.

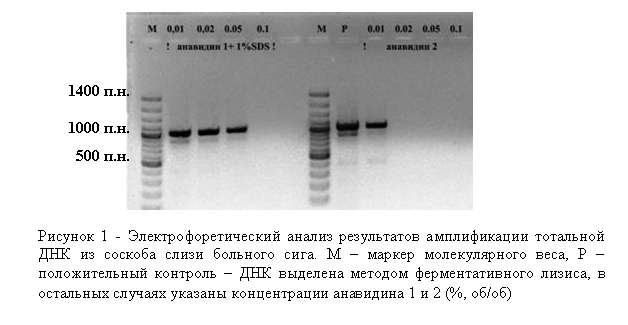
Известно, что этидиумбромид встраивается в двойную цепь ДНК, что способствует ее разрушению при облучении УФ-светом. Установлено, что обработка клеток производным этидиумбромида – моноазидом (ЕМА) с последующим кратковременным освещением светом высокой мощности приводит к селективной инактивации как внеклеточной ДНК, так и ДНК, находящейся в мертвых или частично разрушенных клетках (Nocker, Camper, 2006; Wagner et al., 2008). Механизм этого процесса основан на неспособности этого химического реагента проникать через клеточную стенку жизнеспособных и активных клеток.
При мониторинге бактериальных заболеваний существует необходимость получения достоверной информации: из живых или мертвых бактериальных клеток получен ампликон. Актуальность проведения данной диагностики вызвана следующими особенностями: 1) в пробах, взятых с внешних покровов рыб, особенно с травмированных участков тела, присутствуют как бактериальные, так и мертвые (лизированные) клетки эпителия организма-хозяина; 2) специфика профилактики и лечения бактериальных инфекций, заключается в регулярной обработке аквариумов дезинфицирующими средствами, разрушающими клетки микроорганизмов.
Мы использовали моноазид этидиумбромида для селективной инактивации внеклеточной ДНК, ДНК отмерших эпителиальных клеток организма-хозяина, а также мертвых бактериальных клеток. Следует отметить, что вся процедура дополнительной обработки занимает несколько минут (обработка светом идет в течение 90 сек). В экспериментах использовали как соскобы с внешних покровов рыб, так и фрагменты кишечника (рис. 3). ПЦР-продукты на некоторых специфичных праймерах, в частности на специфичных праймерах на бактерии рода Bacillus, удалось получить только с тех препаратов ДНК, которые были получены после дополнительной обработки ЕМА.
Очевидно, что соотношение ДНК:ЕМА при первичной обработке биологического материала является принципиально важным и должно быть оптимизировано для полного ингибирования ДНК в мертвых клетках. Были проведены предварительные эксперименты по поиску оптимальных концентраций бактериальных клеток для ингибирования ДНК после их разрушения или фиксации на примере культуры грамотрицательных бактерий Escherichia coli. Результаты показали, что фиксация или разрушение клеточной суспензии, титр клеток в которой меньше 1–2×105 кл./мл, и последующая обработка ЕМА приводят к полному ингибированию тотальной ДНК, т. к. не удается получить ПЦР-продукты на праймерах с разным уровнем специфичности. При титре клеток выше 1×106 кл./мл обработка суспензии клеток ЕМА после их лизиса не влияет на получение ПЦР-продукта. 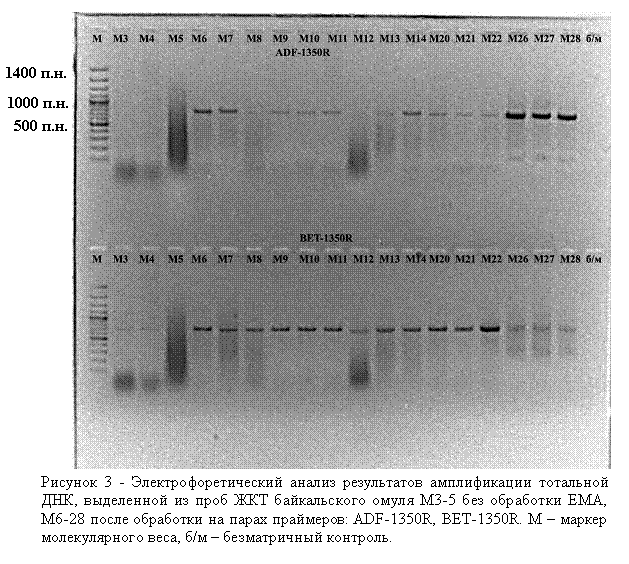
 По-видимому, соотношение ДНК:ЕМА в последнем случае не является оптимальным для полного ингибирования ДНК в мертвых клетках. Это соотношение необходимо учитывать при оценке оптимального количества биомассы материала, которое необходимо отбирать при проведении диагностического молекулярно-генетического анализа. Для этого необходимо провести дополнительные эксперименты по определению оптимальных концентрацией ЕМА при обработке микроорганизмов с разным типом клеточной стенки. Однако несомненно, что данный подход является перспективным для дальнейшего исследования и практических рекомендаций.
По-видимому, соотношение ДНК:ЕМА в последнем случае не является оптимальным для полного ингибирования ДНК в мертвых клетках. Это соотношение необходимо учитывать при оценке оптимального количества биомассы материала, которое необходимо отбирать при проведении диагностического молекулярно-генетического анализа. Для этого необходимо провести дополнительные эксперименты по определению оптимальных концентрацией ЕМА при обработке микроорганизмов с разным типом клеточной стенки. Однако несомненно, что данный подход является перспективным для дальнейшего исследования и практических рекомендаций.
На следующем этапе была проведена диагностика бактериальных инфекций лососевидных рыб, вызываемых Aeromonas hydrophila и Pseudomonas anguilliseptica, в условиях аквариумной экспозиции Байкальского музея ИНЦ СО РАН на основе видо-специфичной амплификации. Эти микроорганизмы широко распространенны в естественных водоемах. В благоприятных условиях рыбы обладают устойчивостью к этим видам и вырабатывают вещества, предупреждающие развитие инфекций. Апробацию метода и подбор условий видо-специфичной амплификации проводили для анализа соскобов с кожных покровов рыб. Положительные ПЦР-продукты были получены только с препаратов ДНК, полученных с внешних покровов рыб с явными проявлениями язвенных заболеваний.
Таким образом, разработана эффективная методика, позволяющая корректно детектировать бактериальные инфекции рыб в условиях искусственного содержания с учетом селективной детекции живых и жизнеспособных бактериальных клеток на основе специфичного ингибирования внеклеточной ДНК.
Работа поддержана инновационным проектом «Тест-системы для диагностики бактериальных инфекций рыб» 2008 г. ИНЦ СО РАН и выполнена в рамках программы РАН №23, подпрограммы 1, проект 23.13 (2009-2010 гг., рук. ).
Библиографический список
1. Абрамова направления технологических исследований ВНИРО // Прикладная биохимия и технология гидробионтов : труды ВНИРО. - М. : Изд-во ВНИРО, 2004. - Т. 143. - С. 9–16.
2. , Андреева в молекулярную экологию микроорганизмов : учебно-методическое пособие. – Ярославль : Изд-во , 2009. – 91 с.
3. , , Ханаева методов молекулярно-генетического анализа для изучения микроорганизмов, ассоциированных с рыбами // Биология внутренних вод. - 2008. - № 2. - С. 91–94.
4. Богерук в России: история и современность // Рыбное хозяйство. - 2005. - № 4. - С. 14–18.
5. , , Васильков рыб и основы рыбоводства. – М. : Колос, 1999. – 456 с.
6. , , Плотников микрофлора рыб различных экологических групп // Известия РАН. Серия биологическая. - 2007. - № 6. - С. 728–737.
7. , , , Копыленко продуктов из гидробионтов // Прикладная биохимия и технология гидробионтов : труды ВНИРО. - М. : Изд-во ВНИРО, 2004. - Т. 143. - С. 42–44.
8. Кузьмина -биохимические основы экзотрофии рыб. - М. : Наука, 2005. – 300 с.
9. Austin B. The bacterial microflora of fish // TheScientificWorldJOURNAL. - 2002. - Vol. 2. - P. 558–572.
10. Bentsink L., Leone G. O., van Beckhoven J. R., van Schijndel H. B., van Gemen B., van der Wolf J. M. Amplification of RNA by NASBA allows direct detection of viable cells of Ralstonia solanacearum in potato // J. Appl. Microbiol. - 2002. - Vol. 93. - P. 647–655.
11. Buller N. B. Bacteria from fish and other aquatic animals: a practical identification manual. – Oxfordshire : CABI publishing, 2004. – 361 p.
12. Burnett S. L., Beuchat L. parison of methods for fluorescent detection of viable, dead, and total Escherichia coli O157:H7 cells in suspensions and on apples using confocal scanning laser microscopy following treatment with sanitizers // Int. J. Food Microbiol. - 2002. - Vol. 74. - P. 37–45.
13. Flekna G., Stefanic P., Wagner M., Smulders F. J. M., Mozina S. S., Hein I. Insufficient differentiation of live and dead Campylobacter jejuni and Listeria monocytogenes cells by ethidium monoazide (EMA) compromises EMA/real-time PCR // Res. Microbiol. - 2007. - Vol. 158. - P. 405–412.
14. Haygood M. G. Light organ symbioses in fishes // Crit. Rev. Microbiol. - 1993. - Vol. 19. - P. 191–216.
15. Haygood M. G., Distel D. L., Herring P. J. Polymerase chain-reaction and 16S-ribosomal-RNA gene-sequences from the luminous bacterial symbionts of 2 deep-sea anglerfishes // J. Mar. Biol. Assoc. U. K. - 1992. - Vol. 72. - P. 149–159.
16. Lee J.-L., Levin R. E. Quantification of total viable bacteria on fish fillets by using ethidium bromide monoazide real-time polymerase chain reaction // Int. J. Food Microbiol. - 2007. - Vol. 118. - P. 312–317.
17. Lee J.-L., Levin R. E. A comparative study of the ability of EMA and PMA to distinguish viable from heat killed mixed bacterial flora from fish fillets // J. Microbiol. Methods. - 2009. - Vol. 76. - P. 93–96.
18. McKillip J. L., Jaykus L. A., Drake M. rRNA stability in heat killed and UV-irradiated enterotoxigenic Staphylococcus aureus and Escherichia coli O157:H7 // Appl. Environ. Microbiol. - 1998. - Vol. 64. - P. 4264–4268.
19. Michel C., Antonio D., Hedrick R. P. Production of viable cultures of Flavobacterium psychrophilum: approach and control // Res. Microbiol. - 1999. - Vol. 150. - P. 351–358.
20. Nocker A., Camper A. K. Selective removal of DNA from dead cells of mixed bacterial communities by use of ethidium monoazide // Appl. Environ. Microbiol. - 2006. - Vol. 72. - P. 1997–2004.
21. Nocker A., Mazza A., Masson L., Camper A. K., Brousseau R. Selective detection of live bacteria combining propidium monoazide sample treatment with microarray technology // J. Microbiol. Methods. - 2009. - Vol. 76. - P. 253–261.
22. Novak J. S., Juneja V. K. Detection of heat injury in Listeria monocytogenes Scott A. // J. Food Prot. - 2001. - Vol. 64. - P. 1739–1743.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 |
