

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
23. Rudi K., Moen B., Dromtorp S. M., Holck A. L. Use of Ethidium Monoazide and PCR in Combination for Quantification of Viable and Dead Cells in Complex Samples // Appl. Environ. Microbiol. - 2005. - Vol. 71, - № 2. - P. 1018–1024.
24. Sambrook J., Fritsch E. F., Maniatis T. Molecular Coning. A laboratory Manual. - Cold Spring Harbor : Cold Spring Harbor Laboratory Press, 1989. - Vol. 2. - 345 p.
25. Sheridan G. E., Masters C. I., Shallcross J. A., MacKey B. M. Detection of mRNA by reverse transcription-PCR as an indicator of viability in Escherichia coli cells // Appl. Environ. Microbiol. - 1998. - Vol. 64. - P. 1313–1318.
26. Spanggaard B., Huber I., Nielsen J., Nielsen T., Appel K. F., Gram L. The microflora of rainbow trout intestine: a comparison of traditional and molecular identification // Aquaculture. - 2000. - Vol. 182. - P. 1–15.
27. Van der Maarel M. J.E. C., Sprenger W., Haanstra R., Forney L. J. Detection of methanogenic archaea in seawater particles and the digestive tract of a marine fish species // FEMS Microbiol. Lett. - 1999. - Vol. 173. - P. 189–194.
28. Wagner A. O., Malin C., Knapp B. A., Illmer P. Removal of free extracellular DNA from environmental samples by ethidium monoazide and propidium monoazide // Appl. Environ. Microbiol. - 2008. - Vol. 74, № 8. - P. 2537–2539.
Глава 4. ПОДХОДЫ К КОМПЬЮТЕРНОМУ МОДЕЛИРОВАНИЮ УГЛЕВОД-БЕЛКОВЫХ ВЗАИМОДЕЙСТВИЙ
Углеводы играют очень важную роль в процессах межклеточного распознавания, проникновения в клетку вирусов, лекарcтвенных веществ и т. п. В последнее время все больше внимания уделяется изучению пространственных механизмов биологического взаимодействия углеводов с природными рецепторами. Понимание механизмов физиологического действия углеводов на молекулярном уровне необходимо для направленного дизайна структур с заданной активностью.
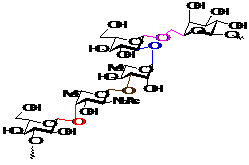
Такая важная роль углеводов определяется особенностями строения их молекул (Рисунок 4). С одной стороны, они являются конформационно подвижными соединениями засчет наличия гибких мостиков между моносахаридными остатками, которые называются гликозидными связями. С другой стороны, сами моносахаридные кольца имеют, как правило, достаточно жесткую конформацию. Однако, наличие многих вариантов взаимного пространственного расположения заместителей в этих остатках обуславливает большую вариабельность пространственных структур углеводов.
Таким образом получается, что молекулы углеводов могут принимать весьма широкий спектр конформаций, в то время как некоторые их фрагменты остаются фиксированными. Это облегчает образование углевод-белкового комплекса и позволяет иногда достичь специфичности определенного углеводного лиганда к определенному белку.
| |
 | |
![]()
![]()

![]()
![]()
![]() Рисунок 4 - Структурные особенности молекул углеводов.
Рисунок 4 - Структурные особенности молекул углеводов.
На настоящий момент существуют различные методики теоретического молекулрного моделирования структур комплексов углеводов с белками, являющимися их природными мишенями. Задачу такого моделирования можно разделить на две части:
1) поиск оптимальной с энергетической точки зрения конформации самой молекулы углевода
2) количественная оценка энергии взаимодействия молекулы лиганда с рецептором.
Методики, позволяющие решать первую часть задачи, известны достаточно давно. Как правило, это методы молекулярной механики, хотя иногда для конформационного анализа применяются и неэмпирические методы квантовой механики.
Для решения второй части существуют так называемые “функции ранжировки” (“scoring functions”), развитие и оптимизация которых и представляет особый интерес. Следует отметить, что не всегда обе части докинга выполняются в одной компьютерной программе. Так, существует ряд программ, позволяющих проводить количественную оценку сродства лиганда к белку, в которых конформации лиганда берутся из сторонних источников, например на основании данных молекулрно-механического расчета или экспериментального конформационного анализа с помощью методов ЯМР. В докладе рассматривается программа XScore [1], в которой реализованы сразу несколько функций ранжировки [2]. Наряду с величинами сродства, вычисленными по каждой из этих функций, программа на выходе предоставляет так их усредненную величину и эмпирически вычисленное значение коэффициента распределения LogP, который так же может оказаться полезным при поиске потенциальных лекарственных веществ.
Так же рассматривается использование программы LigBuilder [3], которая не рассчитывает степень сродства заданного лиганда к белку, но по эмпирическим функциями на основе структуры связывающего центра в белке может предложить структурные модификации лиганда, потенциально приводящие к увеличению активности.
Кроме того, в докладе упоминается возможность самостоятельного построения функции ранжировки с использованием для этого метода молекулярных дескрипторов. Такими дескрипторами могут быть, например, число атомов определенного элемента, молекулярный вес, молекулярный объем и поверхность, дипольный момент, коэффициент распределения и множество других. Смысл данного подхода состоит в том, что строится математическая модель, коррелирующая все эти параметры с некоторой экспериментальной величиной, характеризующей активность лиганда по отношению к белку. Чаще всего в качестве такой величины используется константа 50%-ого ингибирования, IC50. Для построения такой модели могут быть использованы самые разные математические приемы, в том числе, метод искусственных нейронных сетей [4].
В заключение в докладе рассматривается самый современный на сегодняшний день алгоритм докинга, реализующий одновременно как расчет величины внутримолекулярной конформационной энергии лиганда, так и энергии межмолекулярного взаимодействия лиганда с рецептором. В данном случае, следовательно, константа устойчивости получающегося комплекса может быть рассчитана напрямую, а через нее — и величина IC50. Самая распространенная свободно доступная компьютерная программа, использующая данный подход — Autodock [5]. Эта программа имеет следующие преимущества:
1) Точность расчета энергий связывания составляет 2.1 ккал/моль;
2) Можно учитывать эффекты сольватации-десольватации как особый вид взаимодействий;
3) Позволяет учитывать подвижность рецептора.
Так же в докладе отмечено, что с помощью программы Autodock можно проводить анализ вкладов отдельных атомов или других фрагментов молекулы в энергию связывания. Это позволяет предположить исходя из компьютерных расчетов, какие именно из частей лиганда являются существенными для проявления активности.
Таким образом, на сегодняшний день существует ряд функций ранжировки («scoring functions»), позволяющих провести количественную оценку стабильности углевод-белковых комплексов. Эти функции могут быть реализованы в программах, осуществляющих только расчет сродства лиганда к рецептору, в этом случае на выходе получаются эмпирические величины, которые могут быть прокоррелированы с величинами IC50. В случае если программа проводит конформационный анализ лиганда и всего комплекса в целом, на выходе получаются величины изменения свободной энергии образования комплекса, из которых его константы устойчивости и величины IC50 могут быть рассчитаны в явном виде. Анализируя вклады различных частей молекулы в связывание, можно делать предположения о возможном упрощении структуры лиганда с синтетической точки зрения, в том числе, с перспективой создания на его основе неуглеводного потенциального лекарственного вещества.
Библиографический список
1. [Электронный ресурс]. – Режим доступа: http://sw16.im. med. umich. edu/software/xtool/manual/index. html
2. Wang R., Lu Y., Wang parative Evaluation of 11 Scoring Functions for Molecular Docking. // J. Med. Chem. – 2003. - Vol. 46. – P. 2287-2303.
3. [Электронный ресурс]. – Режим доступа: http://mdl. ipc. pku. edu. cn/drug_design/work/ligbuilder. html
4. Fabry-Asztalos L., Andonie R., Collar C. J., Abdul-Wahid S., Salim N. A. Genetic Algorithm Optimized Fuzzy Neural Network Analysis of the Affinity of Inhibitors for HIV-1 Protease. // Bioorg. Med. Chem. – 2008. – Vol. 16. – P. 2903-2911.
5. [Электронный ресурс]. – Режим доступа: http://autodock. scripps. edu
Глава 5. бактериальные биопленки и ремедиация среды
Известно, что микроорганизмы являются наиболее древней формой жизни на Земле. Они же являются и основными преобразователями среды. Если на планете произойдет какая-нибудь глобальная катастрофа, то микроорганизмы – бактерии и вирусы, несомненно, выживут и на основе их генетического материала начнут образовываться новые формы жизни.
То, что бактерии способны к самоорганизации и образуют сообщества, было известно еще с середины минувшего столетия. Но активное изучение этой особенности началось в 90-х годах XX-го столетия, а первые публикации, касающиеся такой формы организации, как биопленка, датируются 2002-2003гг.
Способность бактерий образовывать биоплёнки интересна ввиду того, что микроорганизмы проявляют устойчивость к действию антимикробных веществ при их росте в биоплёнках. Биоплёнки - высокоупорядоченные бактериальные сообщества, которые позволяют бактериям жить в прикреплённом состоянии. Биоплёнки могут состоять из одного или нескольких видов бактерий. Их пронизывает сеть водных каналов, обеспечивающих доставку питательных веществ членам сообщества и удаляющих продукты метаболизма. В одной биоплёнке можно наблюдать различные образцы генной экспрессии, что говорит о том, что индивидуальные члены сообщества имеют «специфические обязанности», которые, комбинируясь с другими, усиливают жизнеспособность всего консорциума. Микроколонии в зрелой биоплёнке расположены во внеклеточном полисахаридном матриксе. Итак, биопленка – это взаимодействующая общность разных типов микроорганизмов, сгруппированных в микроколонии, окруженные защитным матриксом.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 |
