
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Электрический заряд является вторым по значимости фактором, определяющим фильтруемость макромолекул. Для любой исходной величины заряда, отрицательно заряженные макромолекулы фильтруются в меньшей степени, а положительно заряженные макромолекулы — в большей степени, чем электронейтральные молекулы. Дело в том, что поверхность всех компонентов фильтрационного барьера (клеточная эндотелиальная выстилка, базальная мембрана и
43
поверхностный слой на подоците) содержат фиксированные полианионы, которые отталкивают отрицательно заряженные макромолекулы во время процесса фильтрации.
Поскольку подавляющее число белков плазмы несет почти только отрицательные электрические заряды, это препятствие в виде электрического заряда играет очень важную ограничительную роль, увеличивая значимость барьера, который реагирует только на величину молекулы. (Например, когда электронейтральный декстран с величиной молекулы, близкой к величине молекулы альбумина, назначается экспериментальным животным, то оказывается, что они (молекулы декстрана) фильтруются на 5—10 %, т. е. значительно больше, чем соответствующие 0,02 % молекул альбумина). При определенной форме патологии почки, когда почечное тельце становится «проницаемым» по отношению к белкам, это происходит, поскольку на мембранах исчезает отрицательный заряд.
Следует указать, что отрицательный заряд на фильтрирующих мембранах служит препятствием только по отношению к макромолекулам, но не по отношению к неорганическим ионам или низкомолекулярным органическим растворенным веществам.
В конечном итоге следует заметить, что определенные низкомолекулярные растворенные вещества, которые в другой ситуации могут быть профильтрованы полностью, в реальности частично связаны с крупными белками плазмы; та часть молекул, которая будет связана с альбумином, не будет фильтроваться через стенку клубочка.
Концентрация такого вещества в боуменовой капсуле будет равняться не абсолютной концентрации в плазме, но концентрации в плазме вещества, не связанного с белком. Например, 40 % кальция э плазме связано с белком и, таким образом, концентрация кальция в гломерулярном фильтрате составляет 60 % от его концентрации в плазме. (Такие примеры являются основанием для использования слов «большая часть» в первом предложении раздела.)
![]()
Скорость движения жидкости при фильтрации в любом участке капиллярного русла в организме определяется гидравлической проницаемостью стенки капилляров, площадью их поверхности и результирующим фильтрационным давлением (РФД), действующим через стенку капилляров:
скорость фильтрации - гидравлическая проницаемость х площадь поверхности х
х РФД.
Поскольку трудно вычислить площадь поверхности капиллярного русла, то параметр, называемый фильтрационный коэффициент (К^), используется для определения результата взаимодействия гидравлической проницаемости и площади поверхности3. Показатель РФД является алгебраической суммой гидростатического давления и осмотического давления, обусловленного белком — онкотичё-ское давление (или коллоидно-осмотическое) — по обе стороны стенки капилляра, Применяя эти расчеты к гломерулярным капиллярам, получим:
44
РФД= (Рgс+Пвс) (Рвс + Пgс)
силы, способствую - силы, препятствую
щие фильтрации щие фильтрации
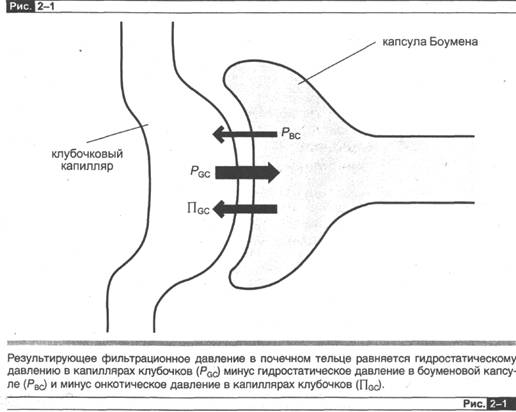
где Рgсс — клубочково-капиллярное гидростатическое давление; Пвс ~~ онкотическое давление жидкости в боуменовой капсуле; РВС — гидростатическое давление в боуменовой капсуле; Пgс ~ онкотическое давление в плазме клубочковых капилляров.
Поскольку белка в боуменовой капсуле практически нет, то величину Пвс можно принять равной нулю и не учитывать в наших расчетах (рис. 2-1). Учитывая сказанное, уравнение для расчета скорости клубочковой фильтрации можно представить таким образом:
СКФ=
Kf
(гидравлическая про-
ницаемость х площадь
поверхности тела)
РФД
(Pgc-Pвс~ Пgс)
Величина гидростатического давления в гломерулярных капиллярах и боумено - вой капсуле у человека не поддается прямым методам измерения. В то же время некоторые результаты непрямых исследований дают возможность предположить,
|
45
46
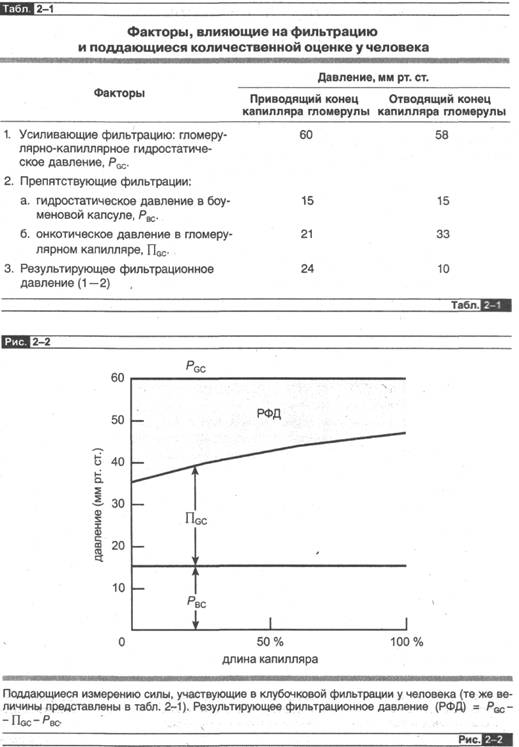
что соответствующие показатели у человека, вероятно, сходны с теми, что получены у собаки. Эти величины представлены в табл. 2-1 и на рис. 2-2 вместе с величинами гломерулярно-капиллярногЪ онкотического давления.
Известно, что гидростатическое давление меняется очень незначительно в различных отделах клубочка; это происходит потому, что самая большая величина поперечной площади сечения гломерулы создает только весьма незначительное сопротивление движению жидкости.
Очень важно обратить внимание, что онкотическое давление в капиллярах гломерулы меняется совсем немного по длине гломерулы; поскольку фильтрат практически лишен белка, то в ходе фильтрационного процесса осуществляется перенос воды, но не белка из плазмы, тем самым увеличивается концентрация белка и отсюда онкотическое давление в непрофильтровавшейся плазме, оставшейся в гломерулярных капиллярах4. В значительной степени из-за этого существенного увеличения онкотического давления чистое фильтрационное давление снижается существенно по направлению от начала к кошту гломерулярных. капилляров, и средняя величина его примерно равна 17 мм рт. ст.5 То, что такой величины давления достаточно, чтобы обеспечить фильтрацию 180 л жидкости в сутки, связано в основном с тем фактом, что гидравлическая проницаемость и, следовательно, К/ в гломерулярных капиллярах на несколько порядков выше, чем во внепочечных капиллярах.
Величина скорости клубочковой фильтрации не является постоянной и может существенно колебаться при различных физиологических состояниях и при патологии. Если все остальные факторы остаются неизменными, любая динамика величин показателей Щ, Рсо Рвс и Пес будет также вести к изменению величины, скорости клубочковой фильтрации. В то же время «все другие факторы» также не остаются постоянно неизменными, и поэтому другие одновременно протекающие события могут противодействовать влиянию специфического фактора, который учитывался в анализе. Фраза «стремится к...» в последующей дискуссии отражает этот факт.
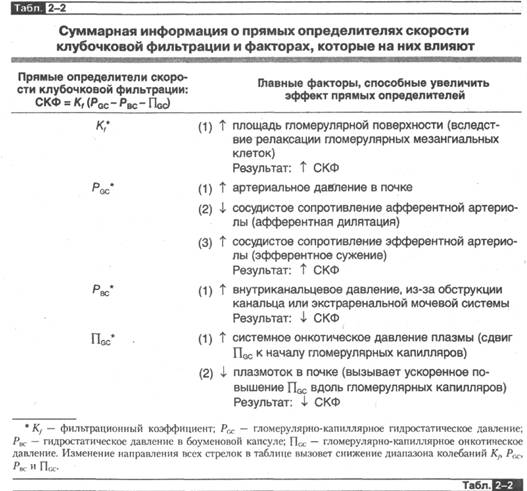
В табл. 2-2 представлена суммарная информация о материале^ описанном в оставшейся части данного раздела. По сути данный материал дает повод для анализа в попытке понять, как патология или вазоактивные химические мессендже-ры и лекарства изменяют скорость клубочковой фильтрации.
В этом контексте следует отметить, что главная причина уменьшения скорости клубочковой фильтрации при патологии почек не изменение указанных параметров в пределах отдельного нефрона, а гораздо более простая — уменьшение числа функционирующих нефронов.
Кf. Изменение Кf может быть вызвано патологией клубочков или лекарствами, но этот показатель также контролируется в ходе обычных физиологических процессов целой серией химических мессенджеров. Механизм неизвестен, но одна вероятная гипотеза заключается в том, что эти мессенджеры вызывают сокращение гломерулярных мезангиальных клеток, что в результате уменьшает площадь гломерулярной поверхности и Кf. Это уменьшение величины Kf влечет за собой снижение СКФ 6.
47 |
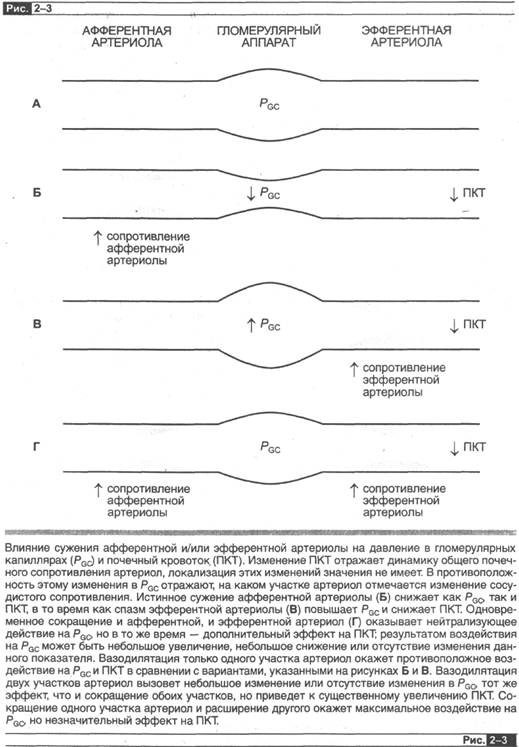
рgс» Клубочково-капиллярное давление отражает взаимодействие почечного артериального давления, сосудистого сопротивления афферентной - артериолы (КА) и сопротивления эфферентной артериолы (КЕ) (рис. 2-3). Первое (не показано на рис. 2-3) изменение почечного артериального давления будет вызывать уменьшение клубочково-капиллярного давления в одноименном направлении (но по соображениям, описанным позднее в этой главе, данная динамика будет минимальной). Второе (рис. 2-ЗБ) при каждом данном значении почечного артериального давления увеличение показателя КА (благодаря спазму афферентной артериолы) будет сопровождаться снижением клубочково-капиллярного давле-ния, просто вызывая большее падение давления между почечными артериями и гломерулярными капиллярами. Напротив, снижение КА (в результате расширения афферентной артериолы) будет весТи к росту клубочково-капиллярного давления. Третье (рис. 2-ЗВ) и более сложное для зрительного восприятия — это на - блюдение, что изменение в КЕ также вызывает динамику клубочково-капилляр-
48
Почечный кровоток и клубочковая фильтрация
|
1ломерулярная фильтрация
49
ного давления, но эти изменения противоположны тем, что вызваны изменением КА. Таким образом, увеличение КЕ (благодаря сокращению эфферентной артерио-лы) вызывает повышение клубочково-капиллярного давления. Так происходит, поскольку эфферентная артериола располагается после гломерулы, так что сужение эфферентной артериолы вызывает задержку крови в гломерулярных капиллярах, увеличивая клубочково-капиллярное давление. Аналогично снижение КЕ (в результате расширения эфферентной артериолы) снижает клубочково-капиллярное давление. Также будет ясно, что когда КА и К. Е будут изменяться одновременно и однонаправленно (т. е. оба увеличиваться или уменьшаться), то это вызывет противоположное воздействие на клубочково-капиллярное давление (рис. 2-ЗГ). Когда они изменяются в противоположных направлениях, они оказывают аддитивное воздействие на клубочково-капиллярное давление.
Теперь давайте вспомним факторы, регулирующие почечный кровоток, и посмотрим, насколько противоположным образом ситуация меняется для данного параметра; для ПК изменение КА и КЕ в одном направлении вызывает аддитивное воздействие на почечный кровоток, в то время как изменение в противоположных направлениях ведет к обратному эффекту.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 |
