
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рассмотрим следующую ситуацию: человек выпивает за короткий промежуток времени 1 л пресной воды без сахара, содержащей незначительное количество натрия или других растворенных веществ. Избыток воды снижает осмолярность жидкостей организма, что рефлекторно тормозит секрецию АДГ через гипотала-мические осморецепторы. В результате водная проницаемость собирательных трубок существенно снижается, очень мало воды реабсорбируется из этих сегментов (или реабсорбция совсем прекращается) и экскретируется большой объем очень разбавленной (гипоосмотичной) мочи. Таким образом, из организма удаляется избыточная вода.
При другой крайней ситуации, когда возникает дефицит воды, допустим, при невозможности поступления ее в организм, осмолярность жидкостей организма увеличивается, рефлекторно стимулируется секреция АДГ, возрастает водная проницаемость собирательных трубок, реабсорбция воды при этом максимальна, экскретируется очень неболыцой объем весьма концентрированной (гиперосмо-тичной) мочи. В результате экскретируется относительно меньше профильтро-
161 |
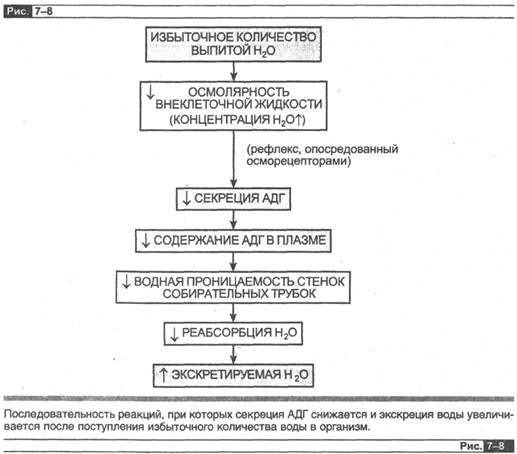
вавшейся воды, чем растворенных веществ (что эквивалентно поступлению чистой воды в организм), и осмолярность жидкостей организма восстанавливается до нормальной.
Теперь мы обрисовали два основных афферентных пути регуляции деятельности гипоталамических клеток, секретирующих АДГ; один, начинающийся от барорецепторов, другой — от осморецепторов. Указанные клетки гипоталамуса интегрируют в одно целое указанные пути, а интенсивность деятельности этих клеток определяется общим количеством информации, переданной через соответствующие синапсы. Из сказанного следует, что одновременное увеличение объема плазмы, и снижение осмолярности жидкости организма вызывает серьезное торможение секреции АДГ. И, соответственно, одновременное снижение объема плазмы и увеличение осмолярности является сильнейшим стимулятором секреции АДГ. Но что происходит, когда импульсы со стороны осморецепторов противоречат импульсам со стороны барорецепторов, например, если объем плазмы и осмолярность снижаются? Обычно из-за большей чувствительности осморецепторов они оказывают доминирующее влияние, когда изменения осмолярности и объема плазмы колеблются от небольших до средних по величине. В то же время очень значительное изменение объема плазмы будет доминировать над снижени-
6 Зак.600
162 Регуляция экскреции натрия и воды: поддержание-постоянства объема плазмы
ем показателя осмолярности жидкостей организма при воздействии на се^срецию АДГ; при этом вода задерживается в большей степени, чем растворенные вещества, и жидкости организма становятся гипоосмотичными (по тем же причинам содержание натрия в плазме снижается).
Здесь еще необходимо добавить, что секретирующие АДГ клетки получают синаптические входы от многих других отделов головного мозга. Из этого следует, что секреция АДГ и, конечно же, мочеотделение могут изменяться под воздействием боли, страха и множества других факторов, включая лекарственные вещества, а также алкоголь, который ингибирует освобождение в кровь АДГ. Эта дополнительная информация, усложняющая картину, не должна затемнить вывод о том, что секреция АДГ определяется в конечном итоге уровнем осмолярности жидкостей организма и объемом плазмы.
Заболевание, которое называется несахарный диабет, отличается от сахарного диабета тем, что возникает вследствие нарушения функционирования системы секреции АДГ. Несахарный диабет характеризуется наличием постоянного водного диуреза (до 25 л/сут). Большинство больных несахарным диабетом утрачивают способность продуцировать АДГ вследствие повреждения гипоталамуса21. Таким образом, водная проницаемость собирательной трубки в данном случае является низкой и не изменяется при увеличении осмолярности и объема внеклеточной жидкости. Другие заболевания вызываются чрезмерно высокой секрецией АДГ. Легко угадать, что у пациентов с патологией такого рода отмечается снижение осмолярности плазмы (и концентрации натрия) вследствие избыточной реаб-сорбции осмотически свободной воды.
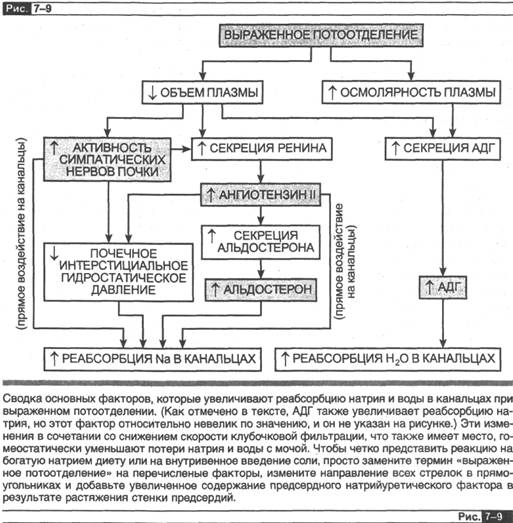
На этом мы завершаем описание регуляции почечной экскреции натрия и воды. На рис. 7-9 суммированы основные факторы, которые участвуют в регуляции данных процессов при реакции организма на обильное потоотделение. Пот является гипоосмотичной жидкостью, содержащей преимущественно воду, натрий и хлор, поэтому потоотделение вызывает и снижение объема внеклеточной жидкости, и увеличение осмолярности жидкостей организма. Задержка почкой натрия и воды помогает компенсировать потери, происходящие при потоотделении.
Жажда и потребность в соли
Теперь мы перейдем к рассмотрению другого компонента водного равновесия, регуляции приема рег оз соли и воды. Следует подчеркнуть, что значительный недостаток соли и воды может быть только частично компенсирован посредством сохранения этих веществ, причем прием их рег оз является конечным компенсаторным механизмом.
Центры, которые опосредуют чувство голода, расположены в гипоталамусе (очень близко к зоне, где продуцируется АДГ). Субъективное ощущение жажды, которое заставляет человека принимать рег оз воду, вызывается как снижением объема плазмы, так и увеличением осмолярности жидкостей организма. Адаптивное значение обоих факторов очевидно. Обратите внимание на то, что упомянутые факторы идентичны тем переменным величинам, которые стимулируют продукцию АДГ, а рецепторы — осморецепторы и барорецепторы сердечно-сосудистой системы, которые инициируют регулирующие продукцию АДГ рефлексы,
163 |
расположены в той же самой зоне, что и центры, отвечающие за жажду. Ощущение жажды воздействует на экскрецию значительно слабее, чем секреция АДГ.
Существуют и другие пути регуляции жажды. Например, сухость во рту и горле вызывает выраженную жажду, которая облегчается при смачивании этих участков слизистой оболочки. Животные, например верблюд (у человека это несколько иначе), при выраженной дегидратации начинают быстро пить, потребляя ровно столько жидкости, сколько необходимо для возмещения потери воды, а затем прекращают пить. Удивительно то, что, когда они останавливаются, вода еще не успевает абсорбироваться из желудочно-кишечного тракта в кровь. В желудочно-кишечном тракте есть определенный механизм, позволяющий измерить объем принятой рег оз воды, но природа его неразгадана.
Ангиотензин II — другой стимулятор жажды, воздействующий непосредственно на мозг; этот гормон стимулирует жажду при снижении объема внеклеточной жидкости.
164_________ Регуляция экскреции натрия и воды: поддержание постоянства объема плазмы
Потребность в соли — аналог чувства жажды — является также чрезвычайно важным компонентом гомеостаза натрия у большинства млекопитающих. Очевидно, что солевой аппетит у млекопитающих является врожденным и складывается из двух крмпонентов: (1) гедонистической и (2) регуляторной потребности. Иначе говоря, (1) животные любят есть соль и потребляют ее при любой возможности, независимо от того, есть ли у них в данный момент дефицит соли или нет, и (2) их тяга к добыванию соли существенно увеличивается при наличии у них в организме дефицита данного вещества.
Значимость этих данных о животных для человека неясна. Желание потреблять соль есть у человека, если у него отмечается выраженный ее недостаток, но такая потребность в соли для ежедневного гомеостаза натрия у здорового человека, вероятно, невелика. С другой стороны, у человека существует гедонистическая потребность в соли, что проявляется почти всеобщим потреблением значительных количеств натрия, поскольку этот продукт недорог и доступен. Среднее потребление соли американцами составляет 10—15 г/сут, и это притом, что человек может существовать совершенно нормально при потреблении 0,5 г/сут. Есть доказательства, хотя и противоречивые, что значительное потребление соли может быть фактором патогенеза артериальной гипертонии у предрасположенных к этому субъектов.
Выводы о физиологических эффектах ангиотензина II
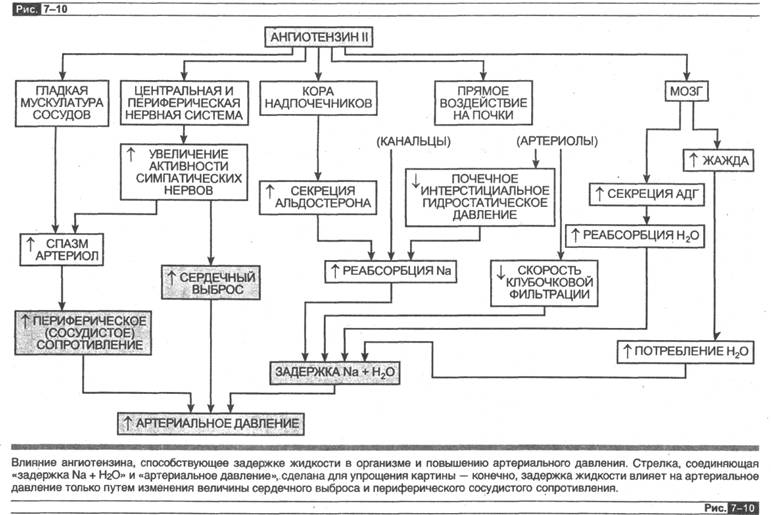
В данной главе и главе 2 мы описали различные физиологические эффекты, вызванные ангиотензином II, и сейчас мы подытожим эту информацию. Ангио-тензин II вызывает разнообразные эффекты в различных частях организма, которые направлены в конечном счете на задержку соли и подъем артериального давления. На рис. 7-2 суммарно указаны эти реакции, кроме того, отмечен факт, ранее не упоминавшийся: ангиотензин II способствует активации симпатической нервной системы (например, посредством увеличения количества норадреналина, освобождаемого в момент потенциала действия). Очень важно понять, что секреция ренина увеличивается вследствие физиологической стимуляции — такой как истощение запасов натрия в организме. Все реакции, показанные на рис. 7-10, направлены на минимизацию водного истощения и снижение показателей артериального давления крови ниже нормы. В противоположность этому, когда соответствующее увеличение секреции ренина возникает в результате болезни (как, например, при стенозе а. гепаНз), эти реакции будут направлены на подъем артериального давления до показателей, превышающих норму.
Вопросы для изучения: 41—56.
Примечания
1 В примерах, приводимых в данном разделе, изменение величины онкотического давления плазмы артериальной крови происходит в направлении восстановления солевого баланса посредством уменьшения или увеличения скорости клубочковой фильтрации и следовательно, экскреции соли. В случае с кровотечением, между тем, этого не происходит;
|
166

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 |
