

Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Показатели | Варианты опыта | |||
ГХЦЖК (контроль) | ГХЦЖК + унитиол+мочевины (опыт) | Разница ± | в % | |
Подвижность, баллов | 3,0±0,02 | 3,1±0,01 | + 0,1 | 3,3 |
Скорость движения, мкМ/с | 95,8±8,05 | 96,2±7,64 | + 0,4 | 0,4 |
Абсолютный показатель живучести, ч | 2,5±0,01 | 3,4±0,02 | + 0,9 | 36,0 |
Время редукции метиленовой сини, мин | 16,7±0,45 | 14,5±2,03 | - 2,2 | -15,2 |
Сохранность SН-групп, % | 62,2±5,13 | 70,4±6,52 | + 8,2 | 13,2 |
Число поврежденных акросом, % | 31,8±3,04 | 23,7±1,86 | - 8,1 | -34,2 |
Активность общих дегидролгеназ, мин | 54,4±6,06 | 48,7±4,63 | - 5,7 | -11,2 |
Активность цитохромоксидазы, мин | 6,8±0,16 | 14,4±0,09 | + 7,6 | 211,7 |
Таким образом изменение состава среды введением унитиола в концентрации 0,028% и мочевины 0,30% позволяет повысить процент подвижных клеток и абсолютный показатель живучести после замораживания-оттаивания. Улучшение функциональную полноценность заморожено-оттаянной спермы хряков дали нам основание включить этих веществ, в состав разрабатываемой новой среды для криокнсервации спермы хряков.
3.3.5 Исследование влияния тиоловых соедиенеий на некоторые структурно-метаболические показатели заморожено-оттаянной сперматозоидов
Для выяснения механизма действия тиоловых соединений на криоустойчивость сперматозоидов и определения направления для дальнейшего совершенствования технологии замораживания было исследовано влияние тиоловых соедиенеий на структурно-метаболические показатели гамет хряков. После замораживания-оттаивания у сперматозоидов оценивали сохранность акросом, ферментов лактатдегидрогеназы и цитохромоксидазы.
Наряду с этим также определяли интенсивность дыхания и состояние дыхательной цепи. Как видно из таблицы 55 унитиол, дитиотреитол и меркаптоэтанол достоверно снижает долю сперматозоидов с поврежденной акросомой.
Таблица 55- Влияние тиоловых соедиений на сохранность акросом после замораживания и оттаивания спермы
Вариант опыта | Число сперматозоидов с поврежденными акросомами, % | ||
M+m | lim | Р | |
ГХЦЖКМ - контроль | 28,42±3,12 | 10,7 - 35,6 | - |
ГХЦЖКМ - унитиол | 20,76±2,56 | 12,9 - 26,8 | 0,05 |
ГХЦЖКМ- меркаптоэтанол | 18,12±3,23 | 5,8 – 25,6 | 0,01 |
ГХЦЖКМ-цистеин | 30,48±4,02 | 13,6 – 40,3 | 0,02 |
ГХЦЖКМ-дитиотреитол | 20,78±3,56 | 11,9 – 26,6 | 0,01 |
При добавлении в среду дитиотреитола и унитиола сохранность акросом сперматозоидов находится на уровне 79,12-79,14%, что выше чес в контроле на 7,8-8,0% (табл. 55).
Анализ полученных результатов (рис. 10) показал, что цистеин не защищает акросому сперматозоидов хряка от повреждающего действия замораживания-оттаивания. Кроме того цистеина не оказывает защитного влияния и на сохранность в сперматозоидах фермента лактатдегидрогеназы (табл. 56). Не отличалась активность лактатдегидрогеназы от контроля и в пробах замороженных с меркантоэтанолом.
Обнаружено также, что по сравнению с контролем унитиол и дитиотреитол достоверно увеличивает сохранность фермента сперматозоидов. Исследование активности цитохромоксидазы измеряемой по времени развития окраски показало, что все тиоловые препараты уменьшают активность данного фермента, участвующего в терминальном участке дыхательной цепи. Наибольшее снижение активности, более 10 раз, наблюдалось в пробах замороженных с меркаптоэтанолом (табл. 56).
Повидимому такое снижение активности (ингибирования) цитохромоксидазы связано с переключением дыхания преимущественно на аэробный гликолиз.
В контрольном опыте было установлено, что используемые в опытах тиоловые соединения обладают гипоксическим действием. Скорость поглощения кислорода из среды унитиолом, дитиотреитолом и меркаптоэтанолом составляет 2...4 натомов О2 в мин.
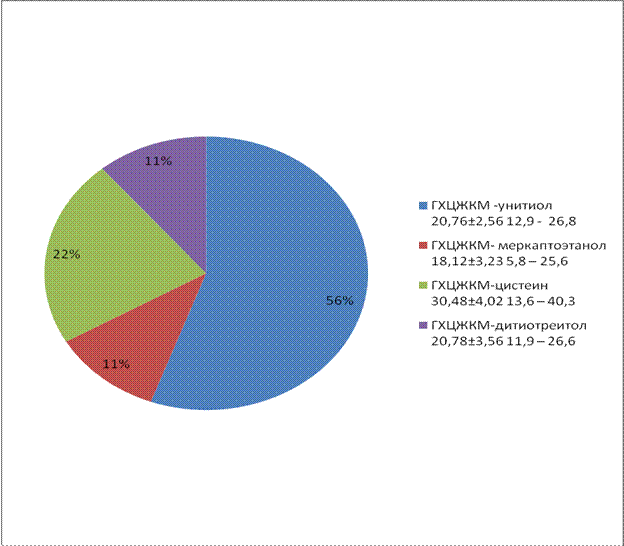
Рисунок 10- Влияние тиоловых соедиений на сохранность акросом
Таблица 56- Активность лактатдегидрогеназы и цитохромоксидазы в замороженно-оттаянной сперме хряков
с тиоловыми соединениями
Вариант опыта | Актиность ЛДГ в мкМ пирувата на 109 сперматозоидов | Активность цитохромоксидазы, мин | ||||
М | D±md | Р | М | D±md | Р | |
ГХЦЖКМ - контроль | 6,9 | - | - | 5,1 | - | - |
ГХЦЖКМ - унитиол | 7,7 | 0,32±0,1 | 0,02 | 15,9 | 10,3±0,9 | 0,05 |
ГХЦЖКМ - меркаптоэтанол | 6,8 | 0,48±0,1 | 0,01 | 54,6 | 48,3±4,1 | 0,01 |
ГХЦЖКМ-цистеин | 7,5 | 0,19±0,1 | 0,02 | 10,9 | 5,8±0,7 | 0,05 |
ГХЦЖКМ-дитиотреитол | 7,6 | 0,31±0,1 | 0,02 | 15,2 | 9,8±0,8 | 0,01 |
3.3.6 Влияние тиоловых соединений на показатель энергетического обмена в сперме хряков на разных этапах замораживания
Энергетические основы подвижности сперматозоидов были разработаны советскими и иностранными учеными (Маnn Т.[34], [167]). Исследователи показали, что центральная роль в энергетике принадлежит аденозинтрифосфату.
Содержание АТФ является важным фактором в регуляции движения сперматозоидов. По данным и др. [90], содержание АТФ в сперме быка и барана 8,7 и 8,6 мг в 100мл, а в сперме хряка - 3,3 мг. Показан параллелизм между содержанием АТФ и подвижностью спермиев. Количество АТФ, быстро расходуемого в процессе движения сперматозоидов, поддерживается на постоянном уровне с помощью двух взаимосвязанных процессов - гликолиза и дыхания. Следует напомнить, что гликолитический процесс слагается из ряда последовательных ферментативных реакций, в результате которых глюкоза оказывается расщепленной на две молекулы молочной кислоты.
Определение АТФ в образцах сперматозоидов. Эксперименты проведены на сперме хряков крупной белой породы. В связи с отсутствием АТФ в семенной плазме, предварительное отделение сперматозоидов от плазмы не проводилось.
Экстрагирование АТФ из сперматозоидов проводилось по методу с соавторами [233]. Для этого из каждого образца спермы отбирали пробу объёмом 0,05 мл и добавляли в неё 0,45 мл диметилсульфоксида. После 20 минутной экстракции при комнатной температуре определяли содержание АТФ биолюминисцентным методом с помощью портативной люминометра ЕМILIТЕ-1003А. Для проведения анализа в кювету люминометра помещали 0,5 мл АТФ реагента, содержащей иммобилизированную люциферазу светляков, люцеферин, соль магния, компоненты буферной смеси и стабилизаторы, и регистрировали фоновое свечение. Затем в кювету вводили экстрагированный образец спермы в объеме 0,1 мл и регистрировали приращение интенсивности свечения (I обр.). Для проведения внутренней калибровки, в ту же кювету вводили 0,1-0,2 мл стандартного раствора АТФ и регистрировали приращение сигнала.
Расчет концентрации АТФ в образце проводили по формуле:
АТФ обр. = Vcт • Jст / Jст • АТФ ст
где: Vст. и Vобр.- объём соответственно стандарта АТФ и образца, АТФ ст.- концентрация АТФ в стандарте.
Содержание АТФ в сперматозоидах животных напрямую коррелирует с их подвижностью и оплодотворяющей способностью ( и др [55], Foulkes J. А. [75], Кichev G. [76], Wilmut I. е. а. [86]).
В связи с совершенствованием методов криоконсервации спермы хряков представляет несомненный практический интерес изучение динамики содержания АТФ в сперматозоидах под влиянием глубокого замораживания в зависимости от методов криоконсервации.
Результаты эксперимента по изучению содержания АТФ в сперматозоидах хряков представлены в табл. 57. Из этой таблицы видно, что в процессе замораживания-оттаивания происходит резкое снижение уровня АТФ в сперматозоидах хряков. Через 5 минут после оттаивания уровень АТФ снизился на 37,7% по сравнению с нативными образцами.
Таблица 57- Влияние глубокого замораживания на содержание АТФ в сперматозоидах хряка
Время исследования | Подвижность сперматозоидов, % | Число сперматозоидов с неповрежденной акросомой, % | Содержание АТФ в сперматозоидах, нмоль/108кл |
|
До замораживания | 81±5,3 | 93±4,2 | 26,8±0,6 |
|
Через 5 мин. после оттаивания | 37±1,9Х | 56±2,8ХХ | 16,7±1,3 |
|
Через 60 мин. после оттаивания | 21±1,3Х | 43+2,9 хх | 7,1±0,3ХХ |
|
Через 120 мин. после оттаивания | 18±0,4ХХ | 37±1,9ХХ | 5,2±0,2ХХ |
|
х-Р<0,05; ХХ-Р<0,01 |
Наряду с этим также уменьшилось на 54,4% подвижность сперматозоидов и 39,8% число сперматозоидов с поврежденной акросомой. Дальнейшая инкубация оттаянных образцов при 37°С способствовала значительному снижению подвижности и содержания АТФ в сперматозоидах. Через 2 часа после оттаивания содержание АТФ в сперматозоидах снизилось в 5,15 раза по сравнению с начальным уровнем (Р<0,01). Следовательно, в процессе замораживания-оттаивания в сперматозоидах хряков происходят значительные деструктивные изменения мембранных структур, в результате чего происходит быстрый гидролиз АТФ под действием ферментов, которые содержатся в клетках и семенной плазме.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 |
