
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
тромба в мелких артериях, артериолах и артериальных капиллярах. В местах, где интенсивность кровотока невелика, роль vWF уменьшается, преобладающим становится взаимодействие, опосредованное другими молекулами, в том числе прямая адгезия тромбоцитов к коллагену посредством GPIa-IIa.
Агрегация тромбоцитов в условиях воздействия активного тока крови тоже происходит с участием фактора Виллебранда. Помимо GPIb-V-IX, с фактором Виллебранда также связывается GPIIb-IIIа. Возможно, что это взаимодействие является ключевым в процессе агрегации в местах сосудистого русла с высокой скоростью тока крови.
Тест агрегации, опосредованный фактором Виллебранда, в лабораторных условиях может быть выполнен с использованием фиксированных тромбоцитов. Видимо, эта реакция не требует энергетических затрат. Однако стимуляция рецептора Ib-V-IX приводит к активации тромбоцита.
Учитывая особенности фактора Виллебранда, можно сказать, что он выполняет функцию «биологического клея», фиксируя тромбоциты на поврежденной сосудистой стенке (рис. 24).
Другая функция фактора Виллебранда - защита ф.VIII от протеолитической деградации системой протеин С - протеин S. В плазме vWF является белком-носителем фактора VIII.
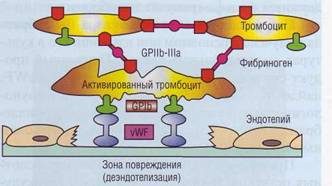
Рис. 24. Фактор Виллебранда (vWF) выполняет роль «биологического клея», прикрепляя к коллагену субэндотелия адгезированные тромбоциты через гликопротеиновый комплекс GPIb-V-IX, Тромб увеличивается в размерах по мере адгезии и агрегации новых тромбоцитов, скрепление которых в агрегат обеспечивает фибриноген, имеющий дива-лентную структуру и взаимодействующий с рецепторами GPIIb-llla
Тромбоциты
Молярная концентрация vWF примерно в 50 раз выше, чем молярная концентрация фактора VIII. Фактор VIII практически весь связан с vWF (рис. 25). Это предупреждает быструю деградацию ф. VШ под влиянием протеина С. Связанный с vWF фактор VIII защищен от протео-литической инактивации в плазме, поскольку у него заблокированы сайты связывания с фос-фолипидной матрицей и заблокированы сайты связывания с протеином С. Поэтому недостаток vWF часто вызывает вторичный дефицит ф.VIII.
В области повреждения сосуда, в процессе vWF-опосредованной адгезии тромбоцитов происходит контакт комплекса vWF-ф. VIII и тромбина (ф. Па), который активирует ф. III, освобождая его из комплекса с фактором Виллебранда.
Фибронектин (плазматический, субэндоте-лиальный и тромбоцитарный) - гранулярный контактный белок, который способен образовывать комплексы с GPIc-Па-рецепторами тромбоцитов и коллагеном. Сродство фибронектина к коллагену и тромбоцитам меньше, чем у фактора Виллебранда, однако молекулярная концентрация его выше. Видимо, фибронектин является основной молекулой адгезии в венозной и капиллярной сети, образуя ось: тромбоцитарный рецептор GPIc-IIa - фибронектин - коллаген. Гликопротеиновый комплекс GPIc-IIa распознает в фибронектине RGD последовательность и осуществляет рецепторную функцию как в интактных, так и в активированных тромбоцитах. Характерная аминокислотная последовательность RGD - трипептид Arg-Gly-Asp имеется во всех адгезивных белках крови, белках а-гранул тромбоцитов, фибриногене, факторе Виллебранда, фибронектине, витронектине и других белках. Наличие RGD-последовательно-сти на фибронектине определяет зависимость процесса его взаимодействия со своим рецептором на тромбоцитах от двухвалентных катионов Са2+ и Mg2+.
Витронектин - гликопротеин плазмы, субэндотелия и а-гранул тромбоцитов. Имеет значение в гемостатических реакциях и в восстановлении поврежденных тканей сосудистой стенки. Витронектин, как и другие адгезивные белки, содержит трипептид RGD, распознающийся интегриновыми рецепторами эндотели-альных клеток и тромбоцитов. Витронектино-
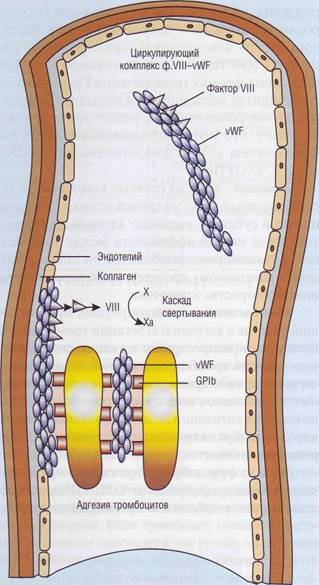
Рис. 25. Комплекс фактор VIII - фактор Виллебранда (ф.Vlll—vWF) состоит из 2 отдельных белков, которые выполняют в гемостазе разные функции, имеют разную химическую и иммунологическую структуру. Фактор VIII необходим для активации фактора X в каскаде свертывания крови, его дефицит вызывает гемофилию А. Фактор Виллебранда (vWF) - полимерный белок, который составляет основную массу комплекса. Он необходим для адгезии тромбоцитов к поврежденной стенке сосудов, обеспечивая взаимодействие коллагена с гликопротеиновым комплексом тромбоцитов GPIb-V-IX. Кроме того, он участвует в агрегации тромбоцитов, взаимодействуя с интегринами GPIIb-llla. Недостаток vWF приводит к болезни Виллебранда
Тромбоциты
вый рецептор на тромбоцитах функционирует постоянно, что отличает его от рецептора фибриногена, который работает только на активированных клетках. У витронектинового рецептора р-цепь аналогична фибриногеновому рецептору (GPIIIa), но а-цепь специфична. При некоторых формах тромбостении Гланцмана на тромбоцитах экспонируется нормальное количество витронектиновых рецепторов, что доказывает, что у этих больных имеет место дефект синтеза а-цепи фибриногенового рецептора, т. е. GPIIb.
Ламинин - один из главных компонентов эк-страцеллюлярного матрикса стенки сосудов, плотный субстрат, вызывает адгезию тромбоцитов. Из-за низкой аффинности между этим белком и рецептором тромбоцитов он лишь содействует адгезивному процессу, причем только при низких скоростях кровотока.
Тромбоспондин - гликопротеин, принимающий участие в адгезии и агрегации тромбоцитов. Он широко распространен в тканях, содержится в α-гранулах тромбоцитов и в небольшом количестве в плазме крови. На поверхности интакт-ных тромбоцитов очень мало тромбоспондина, но после их активации количество экспонированного на мембране тромбоцитов тромбоспондина резко увеличивается.
Одна из функций тромбоспондина - стабилизация комплекса фибриноген-GPIIb-IIIa в процессе агрегации тромбоцитов. Тромбоспондин увели-
чивает его прочность и переводит агрегацию тромбоцитов из обратимой в необратимую (рис. 26).
Помимо этого, тромбоспондин связывается с рядом коагуляционных факторов (тромбином, факторами IХа, Ха), что приводит к повышению их локальной концентрации и защищает от действия ингибиторов.
Активация тромбоцитов
При контакте рецепторов адгезии тромбоцитов с субстратом и под воздействием синтезированного в области повреждения сосуда тромбина начинается процесс активации тромбоцитов. Видимо, основную роль в первичной активации тромбоцитов играет сигнал с рецепторов GPIa-IIa, GPIb-V-IX и GPVI, которые контактируют со своими агонистами, в первую очередь с коллагеном, фактором Виллебранда и тромбином. Помимо коллагена, свойством активировать тромбоциты обладают и другие субэндоте-лиальные структуры.
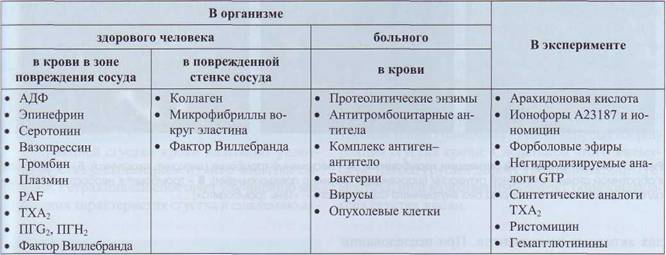
Активация тромбоцитов лежит в основе выполнения ими своих функций. В табл. 5 приведен список основных веществ, активирующих тромбоциты. Почти все эти вещества взаимодействуют с тромбоцитами через специфические рецепторы, которые были описаны выше. Несмотря на многообразие активаторов и большое количество рецепторов к ним, клетка имеет ограниченное количество путей передачи сигнала и эффекторных
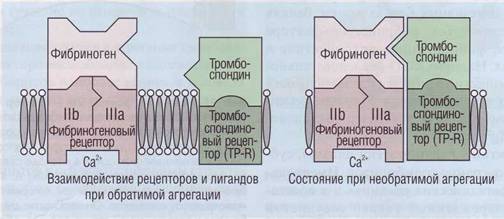
Рис. 26. Взаимодействие рецепторов к фибриногену и тромбоспондину с соответствующими лигандами. При
взаимодействии тромбоцитов с фибриногеном на первой фазе происходит их обратимая активация, При стабилизации комплекса тромбоспондином процесс переходит в необратимую стадию агрегации
Тромбоциты
Субстанции, стимулирующие тромбоциты
Таблица 5
|
Данные приведены по: Kinlough-Rathbone R. L. D. E. MacJntyre, J. L. Gordon. Amsterdam, 1987.
Mustard J. F. // Platelets in biology and pathology, III / Eds
механизмов. Реакция тромбоцита на активирующие воздействия однотипна:
• Тромбоцит меняет форму (рис. 27): у него по
являются псевдоподии, он «распластывается»,
за счет открытой канальцевой системы (ОКС)
увеличивается площадь его поверхности.
• Меняются соотношения различных фосфоли-
пидов между наружным и внутренним лист
ками клеточной мембраны. Это приводит к
появлению на наружной поверхности тром
боцита большого количества кислых фосфо-
липидов с прокоагулянтными свойствами -
фактор 3 тромбоцитов (PF3).
• На мембране тромбоцитов экспрессируются
или повышают аффинность интегрины.
• Происходит секреция содержимого пулов
хранения тромбоцитов во внешнюю среду.
• Тромбоциты фиксируются на поверхностях
(субэндотелиальном матриксе и др.) и (или)
соединяются друг с другом и другими клет
ками крови (происходит адгезия и агрегация).
Активация тромбоцитов может быть обрати мой: происходят лишь частичные конформацион-
ные изменения, обратимое соединение с другими
клетками и частичная секреция гранул. Спустя
небольшое время тромбоцит возвращается в ин-
тактное состояние и поступает в ток крови. Пос
ле обратимой активации и возвращения в неак-
тивное состояние тромбоцит снова может активироваться и вступать во взаимодействие с другими клетками и структурами. Обратимая агрегация возникает при кратковременном воздействии слабого стимула.
Если стимуляция длительная или сильная, происходит необратимая активация тромбоцита. В этом случае тромбоцит прочно фиксируется к другим клеткам или внеклеточным структурам, происходит полная дегрануляция и секреция содержимого пулов хранения. Если тромбоцит после необратимой активации поступает в ток крови, он не может в дальнейшем вступать во взаимодействие с другими клетками и быстро элиминируется из кровообращения. В случае массивного поступления в ток крови необратимо активированных тромбоцитов выявляется достоверное снижение агрегации тромбоцитов со всеми индукторами. Микроскопия в этом случае позволяет выявить большое количество деформированных тромбоцитов.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 |
