
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Плазменные белки гемостаза

Рис. 44. Формирование фибрин-мономеров из фибриногена. Тромбин отщепляет фибринопептиды ФПА и ФПВ от молекулы фибриногена, тем самым образуются растворимые мономеры фибрина, которые способны полимери-зоваться до линейного полимера, или «растворимого фибрина»
створимые мономеры фибрина. В дальнейшем происходит спонтанное соединение комплиментарных участков фибрин-мономеров. Сначала образуются димеры, далее олигомеры и в конечном итоге собираются мононити полимери-зованного фибрина. Таким образом, фибрино-вая цепь формируется спонтанной, конец в конец полимеризацией фибрин-мономеров, при которой концевая часть одного мономера взаимодействует с центральной частью другого мономера в месте отщепления ФПА. Результатом такой полимеризации является линейный полимер шириной в 2 молекулы (рис. 44). На этом этапе фибрин легко растворим в 5-молярной
мочевине, поэтому он получил название растворимого фибрина.
Соединяясь с фибриногеном, тромбин не только отщепляет фибринопептиды. но и активирует связанный с ним фактор XIII. Фактор ХIIIа образует ковалентные связи между γ-цепями (D-доме-нами) нитей растворимого фибрина (рис. 45), которые соединяются за счет образования пептидных мостиков между боковыми радикалами лизина и глютамина. Сшитые между собой мононити фибрина образуют прочную сеть, менее подверженную фибринолизу и более устойчивую к механическим воздействиям. В такой форме фибрин не растворяется в 5-молярной мочевине и называется нерастворимым фибрином.
Рис. 46. Организованный тромб, в котором в фибрино-вую сеть включены клетки крови |
Образовавшийся фибриновый сгусток - трехмерная молекулярная сеть, в которую включены тромбоциты, эритроциты и лейкоциты (рис. 46). Активированные тромбоциты, связанные с нитями фибрина через рецепторы GPIIb-IIIa, сокращают-
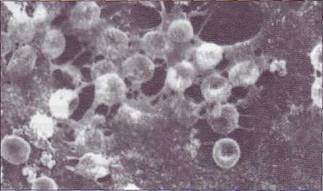

Рис. 45. Образование нерастворимого фибрина под влиянием фактора ХIIIа
Плазменные белки гемостаза
ся под действием тромбостенина (тромбоцитарно-го актомиозина) вследствие присущих им контрак-тильных свойств (см. главу «Тромбоциты»). Происходит ретракция сгустка крови. Сгусток уплотняется, из него выдавливается часть сыворотки. Формирование окончательного тромба наступает на 10-15-й минуте после полимеризации фибрина.
Если тромбоциты отсутствуют или имеют дефект GPIIb-IIIa, то ретракции кровяного сгустка не происходит и он быстро лизируется в процессе фибринолиза. При отсутствии ретракции тромба в сосудистом русле возможен отрыв тром-ботических масс и эмболия удаленных сосудов (тромбоэмболия).
Роль кофакторов и микроокружения в процессе свертывания крови
Плазменные ферменты и ингибиторы системы гемостаза эффективно функционируют только в определенном микроокружении. Эффективность функционирования факторов IX, X, протеина С, антитромбина III в присутствии своих кофакторов (факторы VIII, V, протеин S, гепарансульфат соответственно) возрастает в десятки тысяч раз. Большинство реакций протеолитической активации предшественников ферментов эффективно протекают только на «твердой фазе», роль которой играют отрицательно заряженные фосфоли-пиды. Для всех реакций, протекающих на поверхностях, необходимы ионы кальция. На рис. 47 показана степень активации тромбина фактором Ха в различных условиях микроокружения. Комплекс фактора Ха, фактора Va, фосфолипидов и Са2+ (протромбиназный комплекс) значительно эффективнее активирует тромбин, нежели один фактор Ха, причем в комплексе с кофакторами фактор Ха защищен от деградации антитромбином.
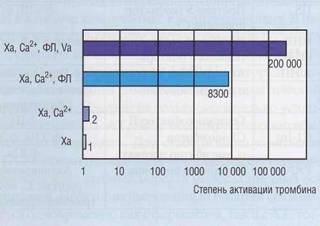
Рис. 47. Влияние условий микроокружения на образование тромбина фактором Ха. Комплекс фактора Ха, фактора Va, фосфолипидов и Са2+ (протромбиназный комплекс) значительно эффективнее активирует тромбин, нежели фактор Ха один или в комбинации только с фосфо-липидами и/или Са2+
Роль кальция в гемостатических реакциях
Роль кальция в гемостазе огромна. Большинство белков гемостаза имеют сайты связывания кальция. При удалении кальция из плазмы (например, при смешивании крови с цитратом натрия) активировать гемостатические реакции практически невозможно. Наиболее важные из известных функций кальция в гемостазе:
• Участие в образовании связей витамин-К-за-
висимых факторов (II, VII, IX, X, протеин С,
протеин S) с фосфолипидной поверхностью.
• Участие в активации фактора XIII.
• Участие в образовании связи ф. VII и ткане
вого фактора.
• Ускорение процесса роста фибринового сгу
стка, участие в стабилизации фибринового
сгустка, ограничение протеолиза фибрина и
фибриногена плазмином, защита фибриноге-
на и фибрина от температурной и щелочной денатурации.
• Стабилизация структуры многих белков ге
мостаза и опосредование взаимодействия
между ними.
• Участие в процессах активации тромбоцитов
и других клеток.
• Кальций необходим для формирования цитос-
келета и возбуждения клетки. Он участвует в
полимеризации актина и миозина и формиро
вании актин-миозиновых волокон. Без него
невозможны процессы изменения формы ак
тивированных клеток, их движение, секреция.
• Кальций участвует в регуляции большинства
внутриклеточных процессов как внутрикле
точный мессенджер (посредник) перемещения
молекул.
Плазменные белки гемостаза
Ингибиторы системы свертывания плазмы крови
Ингибиторы системы свертывания крови представлены в табл. 7.
Ингибиторы системы свертывания плазмы крови
Таблица 7
|
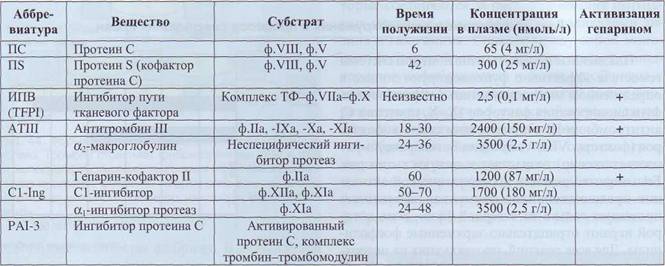
Ингибиторы системы свертывания крови условно можно разделить на три группы - ингибиторы ферментов, ингибиторы коферментов и ингибиторы активных комплексов.
Ингибиторы ферментов системы гемостаза
Среди ингибиторов ферментов системы гемостаза, в свою очередь, можно условно выделить 2 группы - ингибиторы сериновых протеаз и неспецифические ингибиторы протеаз, к которым относится α2-макроглобулин.
Ингибиторы сериновых протеаз, или серпи-ны. Большинство ферментов каскада свертывания крови составляют сериновые протеазы. Се-риновыми протеазами также являются ферменты фибринолитической системы, некоторые ферменты системы комплемента, эластаза, трипсин, химотрипсин и многие другие. Все они имеют гомологичную структуру. Существует группа ингибиторов, специфичных для сериновых протеаз, - серпины. Механизм их ингиби-рующего действия изучен довольно хорошо. Серпины имеют строение, похожее на строение субстрата сериновых протеаз. Однако, охотно соединяясь с ферментами, серпины не подвергаются немедленному расщеплению. Это соеди-
нение блокирует ферментативную активность сериновой протеазы (рис. 48). Различные серпины несколько отличаются по строению, могут быть более или менее специфично ингиби-ровать разные ферменты. Кроме того, на актив-
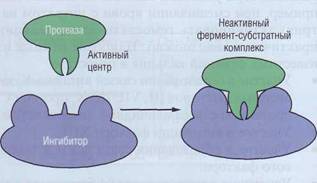
Рис. 48. Ингибирование активных сериновых протеаз серпинами за счет образования стабильного неактивного фермент-субстратного комплекса
Плазменные белки гемостаза
ность и специфичность серпинов может влиять микроокружение.
Антитромбин и гепарин
Антитромбин (синоним - антитромбин III, AT) - гликопротеин, состоит из 432 аминокислот и имеет 4 участка гликолизации с разным количеством сиаловых кислот. Этот ингибитор формирует стабильный 1:1 комплекс с сериновыми протеа-зами плазменного гемостаза. Кроме того, AT связывается со специфическими сульфатными группами на пентасахаридных структурах гепарина.
AT синтезируется в печени и является наиболее значимым ингибитором системы свертывания крови. Активности находящегося в крови здорового человека антитромбина достаточно, чтобы ингиби-ровать в три раза больше тромбина, чем может образоваться из циркулирующего протромбина. Несмотря на это, уже при снижении активности AT в плазме ниже 60% возрастает риск патологических тромбозов. При изолированном дефиците активности AT риск тромботических проявлений возрастает пропорционально степени снижения активности. Помимо тромбина, AT ингибирует фактор Ха, а также факторы IХа, ХIа, ХIIа и калликреин.
Антитромбин по структуре гомологичен α1 - антитрипсину. В его активном центре присутствует специфическая связь Arg-Ser, которая и взаимодействует с сериновыми протеазами.
Рис. 49. Влияние гепарина на активность фактора Ха в плазме. Гепарин существенно усиливает ингибирующий эффект антитромбина на фактор Ха |
Активность AT в десятки тысяч раз усиливается в присутствии отрицательно заряженных гли-козаминогликанов, таких, как гепарансульфат, входящих в структуру гликокаликса на поверхности эндотелиальных клеток. Аналогичное потенциру-
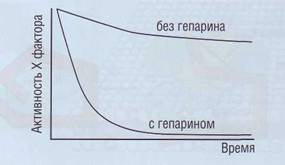
ющее действие на AT оказывает гепарин (рис. 49), вырабатываемый тучными клетками. Антикоагу-лянтное действие гепарина связано с его способностью вызывать конформационные изменения AT. Функция гепарина каталитическая. После образования эквимолярного 1:1 комплекса тромбин-антитромбин (ТАТ) гепарин может освобождаться для организации других комплексов.
Нефракционированный гепарин представляет собой смесь гепаринов различной молекулярной массы. До последнего времени он широко применялся в клинической практике как антикоагулянт. В настоящее время большее распространение получили препараты низкомолекулярного гепарина (НМГ, английская аббревиатура - LMWH), который получается из гепарина химической или энзиматической обработкой. Гепарин не только значительно усиливает активность AT, но и модулирует его ингиби-торную активность. Для стабилизации комплекса ТАТ гепарин должен быть представлен структурой, имеющей, по крайней мере, 18 моносахаридных оснований. Нефракционированный гепарин связывается одновременно как с ферментом, так и с AT, тогда как НМГ связывается только с молекулой AT (рис. 50). Нефракционированный гепарин усиливает активность AT в отношении всех сериновых про-теаз каскада свертывания крови, тогда как низкомолекулярный - в основном в отношении ф. Ха.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 |
