
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Стимуляторы тромбоцитов можно разделить на слабые и сильные.
К слабым стимуляторам относятся АДФ, адреналин, вазопрессин, серотонин. Передача сигнала от рецепторов этих веществ проходит стадию усиления внутри клетки через дополнительный этап образования продуктов тромбоксано-вого завершения и секреции хранимых в грану-
|
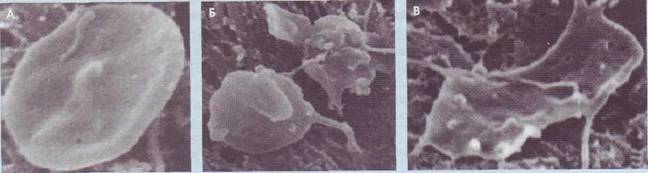
Рис. 27. Стадии контактной активации тромбоцитов: А - неактивный тромбоцит (дискоцит, пластинка); Б - тромбоциты в обратимой стадии контактной активации (шаровидные формы с псевдоподиями); В - тромбоцит в необратимой стадии адгезии (распластанная форма без внутреннего содержимого - «тень тромбоцита»)
лах активных компонентов. При исследовании агрегации тромбоцитов в присутствии слабых стимуляторов на агрегатограммах кривая имеет двухступенчатую форму, что обусловлено усилением агрегации после выделения содержимого пулов хранения (рис. 28).
Сильные стимуляторы тромбоцитов - коллаген, тромбин, большие дозы АДФ - непосредственно после мембранной стимуляции приводят к необратимой активации.
В табл. 5 представлены наиболее важные активаторы тромбоцитов. Часть из них присутствует в подпороговых концентрациях в интактной плазме и избирательно накапливается в зоне повреждения сосудов; другие появляются в системе циркуляции при активации системы свертывания крови в физиологических или патологических условиях. Некоторые факторы выделяются из самих тромбоцитов (АДФ, серотонин, адреналино-подобные субстанции, фактор Виллебранда).
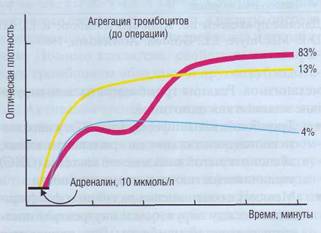
Рис. 28. Типы агрегатограмм. V пациентов при стимуляции агрегации адреналином в дозе 10 мкмоль/л в 83% случаев наблюдается двухфазная агрегация тромбоцитов, в 13% случаев - необратимая агрегация и в 4% - после начальной агрегации наблюдается дезагрегация (собственные данные)
Агрегация тромбоцитов
Процесс агрегации заключается в присоединении активированных тромбоцитов, находящихся в токе крови, друг к другу и ранее фиксированным в области повреждения. Основным рецептором агрегации является GPIIb-IIIa (интег-рин αIIbβ3). После активации тромбоцита GPIIb-IIIa значительно повышает свою аффинность по отношению к фибрину и меняет антигенную структуру (что свидетельствует о значительных кон-
формационных изменениях). После этого происходит соединение тромбоцитов, опосредованное фибрином и фактором Виллебранда (рис. 24).
Вследствие распространения активирующего сигнала на агрегированные тромбоциты, удаленные от места повреждения, образуется толстый слой тромбоцитов, армированный фибрином. Этот процесс лежит в основе образования тром-боцитарного тромба. По мере удаления от зоны
Тромбоциты
повреждения концентрация агонистов активации и агрегации тромбоцитов снижается и соответственно уменьшается активация тромбоцитов. Дистально расположенные частично активированные тромбоциты отрываются от сгустка и возвращаются в кровоток. Таким образом, периферическая дезагрегация тромбоцитов предотвращает неограниченный рост сгустка.
Ретракция сгустка крови
Ретракцией сгустка крови называют уплотнение сгустка с выделением из него избытка сыворотки. Ретракция способствует улучшению механических характеристик сгустка и снижению ак-
тивности фибринолиза внутри него. Ретракция сгустка связана с контрактильными свойствами тромбоцитов. Фибриллы миозина, расположенные в цитоплазме тромбоцитов, фиксированы к мембранному гликопротеину GPIIb-IIIa. В активированных тромбоцитах за счет миозина происходит процесс постепенного «сжимания» цитоплазмы, что приводит к уплотнению всего сгустка крови.
При врожденной недостаточности GPIIb-IIIa - тромбастении Гланцмана - грубо нарушается ретракция сгустка крови. Следствием этого является не только грубый дефект тромбоцитарного гемостаза, но и качественный дефект образовавшегося сгустка крови.
Роль лейкоцитов в гемостазе
РОЛЬ ЛЕЙКОЦИТОВ В ГЕМОСТАЗЕ
Лейкоциты (нейтрофилы и моноциты) в зоне повреждения сосуда участвуют в гемостатических реакциях.
Участие нейтрофилов в пристеночном тромбообразовании
Агрегация тромбоцитов сопровождается освобождением из α-гранул активаторного рецептора Р-селектина (CD62), который остается ассоциированным с плазматической мембраной тромбоцитов. Экспрессия на мембране лейкоцитов Р-селек-тин-связывающего гликопротеина-1 (PSGL-1) позволяет нейтрофилам присоединять тромбоциты (рис. 29). Связь нейтрофилов с тромбоцитами обеспечивает репаративные и воспалительные реакции, возникающие в ответ на повреждение.
Нейтрофилы после связывания на мембранах способны секретировать адгезивные молекулы и интерлейкины. Некоторые из интерлей-кинов, в частности интерлейкин-1 (ИЛ-1) и фактор некроза опухоли-ос (ФНО-а), активируют эндотелиальные клетки. Первичный контакт гранулоцитов приводит к перемещению их вдоль сосудистой стенки с последующей транс-эндотелиальной миграцией в субэндотелий. При действии повреждающих факторов, таких, как иммунные комплексы, эндотоксин, гранулоци-
ты могут дегранулироваться и освобождать ИЛ-1, ФНО-α, протеолитические ферменты, такие, как эластаза и катепсин, активные формы кислорода (О2-, О2+), что в свою очередь ведет к повреждению сосудистой стенки. Этот процесс доминирует при воспалительных реакциях. Протеолитические ферменты, которые освобождаются из лейкоцитов, в участках воспаления вызывают нарушения структуры и функции эндотелия, это является условием развития петехий.
Роль нейтрофилов в модуляции реакций гемостаза требует уточнения. С одной стороны, экспрессия тканевого фактора на мембране нейтрофилов происходит либо при длительной стимуляции различными провоспалительными цитокинами, либо после длительного взаимодействия с Р-селектином активированных тромбоцитов. К этому времени на активированных тромбоцитах уже образуется сгусток крови. С другой стороны, экспериментально доказано моду-
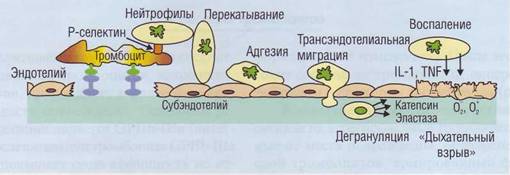
Рис. 29. Участие активных нейтрофилов в повреждении сосудистого эндотелия
Роль лейкоцитов в гемостазе
лирующее воздействие лейкоцитов крови на функцию тромбоцитов при исследовании агрегации тромбоцитов в цельной крови. Кроме
того, исследования показали возможность сборки на нейтрофилах протромбиназного комплекса.
В последнее время описан феномен агрегации лейкоцитов (нейтрофилов) при ишемии тканей. Этот феномен особенно значим для повреждения легких при шоке. В развитии геморрагического шока он играет ведущую роль. На рис. 30 представлен агрегат из нейтрофилов, сформированный на поверхности сосудистой стенки.

Рис. 30. Агрегат из нейтрофилов, выделенный из сосудов легких, в которых формируются агрегаты при развитии респираторного дистресс-синдрома (РДС). Сканирующая электронная микроскопия
Участие моноцитов в свертывании крови
Уникальными свойствами обладают моноци-ты. Это единственные клетки, способные создавать на своей поверхности условия для сборки и успешного функционирования всех ферментатив-ных комплексов системы свертывания крови. Стимулированные моноциты экспрессируют около 16 000 сайтов связывания протромбиназного комплекса. Эффективность синтеза тромбина на их поверхности сопоставима с эффективностью синтеза тромбина на поверхности активированных тромбоцитов.
Синтез и экспрессия тканевого фактора, эффективно связывающего ф. VIIа, происходит на моноцитах под воздействием различных физиологических и патологических стимулов, в том числе бактериальных липополисахаридов, фактора некроза опухоли, интерлейкина-1, С-реактив-ного белка, иммунных комплексов. Сборка теназ-ного комплекса на моноцитарной мембране - ключевой момент в развитии процесса коагуляции. Комплекс тканевой фактор - ф. VIIа подавляется ингибитором пути тканевого фактора (ингибитором внутреннего пути - ИВП), также синтезируемым и экспрессируемым моноцитами.
Реакции свертывания крови, протекающие на моноцитарной мембране, усиливаются специфическими для моноцитов механизмами. Фиксированные на поверхности моноцитов эластаза и ка-тепсин G активируют ф. V до ф. Vа, поэтому ф. V не поступает в кровоток, а остается тут же на мембране моноцитов и формирует протромбиназный
комплекс с ф. Ха. Причем этот комплекс защищен от протеолиза активированным протеином С (АПС), поэтому активность протромбиназы на моноцитарной мембране длительно сохраняется на высоком уровне. Помимо ф. Vа, катепсин G активирует ф. Х. В отличие от катепсина G действие эластазы дозозависимо. В малых концентрациях она активирует ф. V, а в больших - расщепляет ф. Vа. Эластаза, видимо, не обладает способностью инактивировать ф. Ха. Однако воздействие ее высоких концентраций на ф. Х изменяет последний так, что его в дальнейшем невозможно активировать.
Другим альтернативным путем, специфичным для моноцитов, является активация ф. Х после его соединения с мембранным рецептором Мас-1 (CDllb/CD18). Связавшись с Мас-1, ф. Ха частично активирует моноциты и вызывает экспрессию специфического моноцитарного рецептора EPR-1. Комплекс ф. Ха-EPR-l способен эффективно активировать протромбин в присутствии Са2+ без участия ф. Уа. Таким образом, на поверхности моноцита может собираться полноценный протромбиназный комплекс.
Прокоагулянтная активность моноцитов зависит от их микроокружения. Коллагены I и IV типов, фибронектин - активные субстраты для адгезии моноцитов в отличие от ламинина. Последний адгезирует моноциты в 6-10 раз хуже. Однако именно на моноцитах, адгезированных к ламинину, процессы коагуляции развиваются

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 |
