
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Обобщение модели Гаузе для n конкурирующих видов предложено К. Стробеком (Strobeck, 1973) и Д. Логофетом и Ю. Свирежевым (1975):
![]() .
.
К. Стробек исследовал эту систему уравнений на устойчивость и получил, что при n>2 для устойчивости точки равновесия кроме условия αijαji<1, необходимо выполнение дополнительных условий, которые записываются в виде системы 2(n-1) неравенств, зависящих от параметров конкуренции αij и параметров популяционного роста εi. Таким образом, с ростом числа конкурентов сообществу все труднее достигать равновесного состояния.
Другое обобщение модели Гаузе для двух видов предложили Г. Хатчинсон (Hutchinson, 1947, 1948) и У. Каннингам (Canningham, 1955), полагая коэффициенты конкуренции зависящими от численностей видов. Задавая свойства функций в правых частях уравнений подобно тому, как это делал , У. Каннингам получил условия существования устойчивого узла в положительной области.
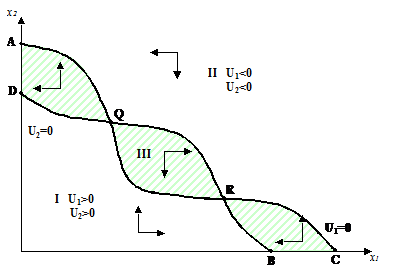
Рисунок 1.15. Обобщенная модель конкуренции двух видов – положение изоклин, при котором существуют две ненулевые точки равновесия в положительном квадранте, причем R - точка устойчивого, а Q - неустойчивого равновесия (по Rescigno and Richardson, 1967).
Колмогоровский подход к исследованию моделей конкуренции применен и в других работах (Rescigno and Richardson, 1967). Изоклины в общем случае описываются не прямыми линиями, и потому могут иметь несколько точек пересечения, которые соответствуют состояниям равновесия. Система уравнений совпадает с приведенной выше моделью хищник - жертва (1-19), но свойства функций в правых частях уравнений иные, чем для системы хищник-жертва:
(1-31)
В примере, приведенном для n =2 (рис. 1-15), изоклины пересекаются в двух точках, и фазовая плоскость делится на три области. Стрелками показаны знаки функций U1 и U2 в каждой из трех областей. Опираясь на заданные свойства функций конкуренции, авторы показывают, что интегральные кривые, начинающиеся в I и II областях, обязательно входят в заштрихованную область III, ограниченную двумя изоклинами. После этого из области DQA интегральные кривые попадут в точку D, а из областей QR и RBC –в точку R. То есть точка Q является точкой неустойчивого равновесия, а точки D и R– устойчивого. Но точка D соответствует случаю, когда первый вид вытесняется вторым.
Если изоклины не пересекаются в положительном квадранте, то выживает только один вид. При B>C (точки B и C – точки пересечения изоклин с осью x1) выживет первый вид, при B<C – второй. Если изоклины пересекаются только в одной точке, то при B<C выживут оба вида, а при B>C – только один, а какой именно - зависит от начальных условий. Для модели Гаузе изоклины Ui=0 являются прямыми линиями и могут пересечься только в одной точке, т. е. модель конкуренции двух видов Гаузе (1-27) является частным случаем модели Ричардсона и Рисайо (1-31).
Модель конкуренции трех видов (Rescigno, 1968) тоже описывается системой (1-31), но при n=3. Функции Ui задаются неявно, автор ограничивается описанием их свойств, но ограничения, наложенные на их свойства, слабее, чем при описании модели (1.31). Исследование показало, что при различном поведении этих функций конкуренция может привести к различным результатам, среди них возможно существование всех трех видов, причем при разных условиях точка равновесия может быть устойчивым узлом, устойчивым фокусом или даже предельным циклом.
Если в описанных выше моделях конкуренции лимитирующий фактор не рассматривается в явном виде, то в модели, предложенной и исследованной нами (Булгакова, 1968а), конкуренция между двумя потребителями ведется за корм, количество которого обозначается через x(t) и пополняется за счет постоянного притока С>0. Система уравнений модели имеет вид:
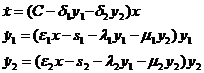 (1-32)
(1-32)
Коэффициенты λ1 и μ2 характеризуют внутривидовую конкуренцию (например, конкуренцию за пищу, каннибализм), а λ2 и μ1- межвидовую конкуренцию.
Приравнивая нулю правые части уравнений (1-32), определяем координаты стационарной точки:
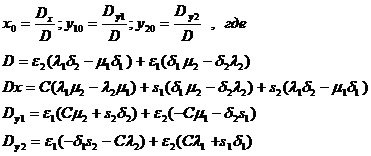
Условия существования положительной стационарной точки имеют вид:
a) min{D, Dx, Dy1,Dy2}>0
или b) max{D, Dx, Dy1,Dy2}<0.
Исследование системы (1-32) на устойчивость решения показало, что необходимое и достаточное условие устойчивости точки равновесия записывается в виде двух следующих неравенств:
min{D, Dx, Dy1, Dy2}>0 и a1a2>a3.
Параметры a1, a2 и a3 представляют собой коэффициенты характеристического уравнения линеаризованной системы и выражаются следующим образом через параметры модели:
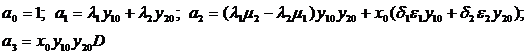
Отметим в частности, что при отсутствии внутривидовой конкуренции (λ1=μ2=0) определитель D<0, т. е. система неустойчива. Таким образом, существование внутривидовой конкуренции является необходимым условием устойчивости равновесия моделируемой системы.
Численность каждого из видов в отсутствие второго вида асимптотически стремится к стационарной точке с координатами x0=(Cλ+δs)/δε); y0=C/δi..
Метод изоклин был применен и при моделировании других взаимоотношений между двумя видами – симбиоза и комменсализма (Gause and Witt, 1935). Для случая симбиоза система уравнений имеет вид:
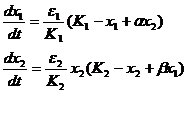 (1-32)
(1-32)
Коэффициенты α и β характеризуют положительное влияние одного вида на другой. Как и в модели конкуренции, изоклины имеют вид прямых линий, но расположены иначе (рис. 1-16).

Рисунок 1.16. Расположение изоклин при симбиозе двух видов
При отсутствии отношений между видами (α = β =0) каждый из них имеет устойчивую точку равновесия, соответствующую пунктирным линиям. При увеличении параметров α и β тангенс угла наклона каждой изоклины растет, и увеличиваются координаты точки равновесия (в пределе эта точка стремится к бесконечности). Точка равновесия является устойчивым узлом с координатами, лежащими в положительном квадранте: 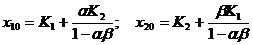 .
.
Если оба параметра α и β отрицательны, получаем рассмотренный выше случай конкуренции. Если один из этих параметров (например, β) отрицателен, а второй положителен, получаем случай комменсализма1,[3]когда второй вид ускоряет рост первого, а первый мешает росту второго (рис. 1-17). При увеличении β точка пересечения изоклин (устойчивый узел) опускается, и в пределе останется только первый вид, а его численность будет равна логистическому пределу роста K1.

Рисунок 1-17. Расположение изоклин при комменсализме двух видов
1.4. Основные выводы главы 1.
Исследование аналитических моделей сообществ на устойчивость проводилось по первой методе (1950) и показало следующее:
1. Простейшая модель Вольтера (Volterra, 1931) имеет точку равновесия только при четном количестве звеньев, причем эта точка находится на границе устойчивости, решение такой системы имеет вид незатухающих колебаний.
2. Автором предложена модель сообщества, в котором запас пищи для организмов первого трофического уровня ограничен (например, количество биогенов в морской системе или количество гумуса в почве для наземного сообщества). Показано, что обратная связь потоков вещества с каждого трофического уровня на нижнее звено стабилизирует динамику сообщества, и устойчивое равновесие может существовать при любом количестве видов. Стабилизирует динамику сообщества и ограниченный приток вещества (или солнечной энергии), необходимого для продуцентов.
3. Исследовано влияние временной задержки τ – длительности времени разложение вещества, поступающего со всех трофических уровней на косную компоненту, до биогенов, потребляемых первым трофическим уровнем. При введении задержки такого типа существует устойчивая точка равновесия, при этом динамика биомассы каждого уровня представляет собой затухающие колебания с задержкой по фазе относительно аналогичного варианта расчетов без задержки.
4. Введение в модель «хищник-жертва» насыщения хищника (рацион которого зависит от количества пищи) делает модель более близкой к реальности. Стационарная точка такой модели устойчива только при учете существования внутривидовой конкуренции вида жертв.
5. Исследованы на устойчивость несколько моделей конкуренции видов. Если в модели конкуренции В. Вольтерра может долго существовать только один вид, в модели конкурирующие виды могут сосуществовать достаточно долго, если экологические ниши этих видов перекрываются не полностью. В модели необходимым условием устойчивого существования двух видов является внутривидовая конкуренция, при этом найдены достаточные условия их сосуществования, представляющие собой неравества функций от параметров системы.
6. Проведен анализ ряда обобщенных моделей взаимодействующих видов, в которых правые части уравнений заданы в виде неявных функций. Впервые такие модели рассматривал (Kolmogoroff, 1936; Колмогоров, 1972). Различные свойства этих функций обеспечивает разные типы взаимодействия видов.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
