
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Понятие "Ответственное рыболовство" введено в 1995 г. Кодексом ФАO по правилам ответственного рыболовства (FAO, 1995). Оно означает, что все организации и отдельные персоны, работающие в области рыболовства, несмотря на разные их роли, должны действовать таким образом, чтобы не наносить ущерб природным ресурсам настоящих и последующих поколений. Все эти субъекты морально, но не юридически, ответственны за свои действия (или за отсутствие действий), если они ведут к исчезновению запасов.
Как видно из приведенных выше определений, эти два понятия практически совпадают. Но это скорее не определения цели, а формулировка необходимых условий, ограничивающих эксплуатацию запасов, причем довольно общая. Для того, чтобы выработать политику регулирования промысла в конкретной экосистеме, которая отвечала бы этим принципам, но и позволяла бы получать оптимальный (по выбранному критерию) вылов разных видов ЭС, необходим экосистемный подход, основанный на многовидовом моделировании. Несомненно, необходимо соблюдать определенные выше принципы, но трудность состоит в том, как реализовать на практике такое регулирование.
Рассмотрим условие сохранения структуры экосистемы. В начале нашей работы утверждалось, что структура ЭС нарушена, если одна из ее компонент исчезает, т. е. ее численность или биомасса стремятся к нулю. Но оказалось, что можно заметить изменения в структуре ЭС еще до того, как она будет близка к разрушению, если проводить мониторинг за некоторыми показателями ее структуры (индексами или индикаторами). В качестве таких показателей М. Синклер и др. (Sinclair et al., 1999) предложили использовать следующие:
- форма размерного спектра, полученного с помощью исследовательских съемок,
- индекс изменения структуры трофических уровней,
- суммарный вылов, получаемый с каждого трофического уровня всеми промыслами.
Эти индексы могут стать основой для определения БО регулирования на уровне экосистемы.
7.4.2. Оценки биологических ориентиров регулирования многовидового промысла
Введение биологических ориентиров управления (БО) позволяет включить в схему управления промыслом неопределенность оценок биомассы и коэффициентов промысловой смертности, ограничивая область допустимых значений управляющих параметров. При одновидовом подходе определяются два БО по биомассе (граничный и предосторожный) и два по промысловой смертности. Как показано на простых примерах в гл.2, для того чтобы при регулировании промысла учесть многовидовые отношения в сообществе, точечные БО по промысловой смертности следует заменять на границы допустимой области регулирования. Определение этих границ в многомерном случае оказывается довольно сложной задачей.
Что же касается БО по биомассе, то на первый взгляд граничные ориентиры можно определять для каждого вида сообщества отдельно, как для изолированных видов, поскольку эти параметры оцениваются из соображений сохранения воспроизводства видов. Но на самом деле для вида, который является кормом для хищников, следует увеличить величину граничного ориентира, поскольку соответствующая ему величина биомассы должна обеспечить не только воспроизводство этого вида, но и поддержать существование и воспроизводство популяций хищников. Предосторожный БО по биомассе вида жертв тоже должен быть увеличен, поскольку к неопределенности оценки его собственных популяционных параметров следует добавить возможную изменчивость популяций хищников. Таким образом, оценивание БО по биомассе предствавляет собой весьма непростую задачу.
На рабочей группе ИКЕС по многовидовым оценкам (ICES, 1997b; Rice, 1997) было исследовано влияние многовидовых отношений типа хищничества и каннибализма на БО, принятые в одновидовом анализе. С этой целью построены имитационные модели, очень упрощенно моделирующие многовидовые отношения. Показано, что пренебрежение многовидовыми отношениями приводит к занижению зависимости Y/R (равновесного улова на единицу пополнения) и завышению зависимости SSB/R (нерестовой биомассы на единицу пополнения) от коэффициента промысловой смертности F, и искажения эти зависят от биомассы хищников. При стабильном промысле эти два искажения могут компенсировать друг друга, и искажение зависимости равновесного улова от F (зависимости Y0(F)) будет невелико. Однако, при нестабильном промысле одновидовой подход может привести к переоценке величины допустимого улова.
Детальное описание построения моделей "Улов (выгрузки) на пополнение", "Выбросы на пополнение" и "Биомасса нерестового запаса на пополнение" приведено С. Муравски с соавторами (Murawski et al., 1991).
Хотя теория определения БО сообщества пока детально не разработана, но при регулировании промысла балтийского шпрота уже принято строить разные зависимости Y/R и SSB/R при разной численности запаса хищника-трески, т. е. разной численности запаса трески соответствуют различные значения ориентиров управления по промысловой смертности шпрота – F01, FMSY и др.
Возможный подход к оценке граничных БО по промысловой смертности показан в гл. 2 на моделях двухвидового сообщества: область допустимого управления на плоскости с координатами {F1,F2} в простейшем случае ограничена прямыми линиями, и вместо точечного БО, принятого в одновидовом случае, граничный БО имеет вид прямой линии на плоскости. В более сложной модели двух видов роль граничного ориентира будет играть кривая линия на той же плоскости, в модели сообщества трех видов – плоскость или поверхность, а в модели n видов – соответственно – n-мерная область.
Х. Гисласон (Gislason, 1999) исследовал влияние межвидовых отношений на величины целевых БО по промысловой смертности (FMSY и F01) и предосторожных БО по биомассе (Bpa и 50% от уровня девственной биомассы) на примере сообщества рыб Балтийского моря. Моделировалось сообщество трех видов – треска, салака (балтийская сельдь) и шпрот – один хищник и два вида жертв, в модель включен также каннибализм трески. Промысел осуществляется двумя типами промыслового усилия – пелагического (совместный промысел салаки и шпрота) и трескового промысла. Проведено сравнение результатов моделирования этого сообщества, полученных по трем когортным моделям – одновидовой VPA, MSVPA и расширению модели MSVPA, обозначенного как MSGVPA. В последней модели рост особей и темп полового созревания хищника-трески моделируются как функции количества доступной пищи. Расчеты по всем моделям проводились с шагом один год.
По каждой модели сначала рассчитывались величины запасов каждого вида в ретроспективе и распределение коэффициентов промысловой смертности F(y, a) по годам и возрастам, затем усреднением функций F(y, a) за несколько последних лет было получено их "статус-кво" распределение по возрастам [F(a)st-quo]. В разных прогонах прогностической модели на ее вход подавалась функция F(a)st-quo, умноженная на различные числа (множители), т. е. в каждом прогоне расчеты для прогнозных лет велись при постоянном распределении промысловой смертности по возрастам. Так были получены долгосрочные равновесные уловы и значения биомассы запасов при разных уровнях (множителях) промысловой смертности.
Влияние межвидовых отношений на целевые БО по промысловой смертности показано на рис. 7.17. Промысловое усилие приводится в относительных величинах по отношению к величине усилия при состоянии системы "статус-кво". Данные расчеты на прогнозный период проведены при условии, что система запас-промысел находится в равновесном состоянии. Промысловое усилие на промысле трески (Ecod), при котором достигается ее максимальный вылов, зависит от усилия пелагического промысла (рис.7.17а, соответствующий критерию FMSY). В расчетах по одновидовой модели VPA максимальному вылову трески соответствует некоторое постоянное промысловое усилие, а в многовидовом случае с ростом усилия пелагического промысла уменьшается биомасса сельди и шпрота, т. е. уменьшается количество доступной пищи для хищников, а значит, усиливается каннибализм трески. Это означает, что увеличивается естественная смертность трески М2. Для получения максимального вылова трески нужно увеличить ее промысловую смертность, при этом уменьшится биомасса взрослых рыб, т. е. и уменьшится каннибализм.
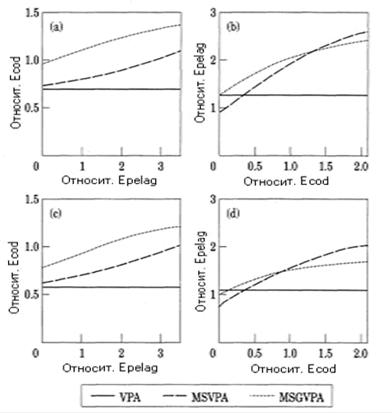
Рисунок 7.17. Зависимость относительного промыслового усилия трескового промысла Ecod, при котором достигается максимальный вылов трески (по критериям FMSY (рис. a) или F01(рис. c)), от относительного усилия пелагического промысла Epelag, а также зависимость относительного усилия Epelag, при котором достигается максимум суммарного улова сельди и шпрота (соответственно критериям FMSY (рис. b) или F01 (рис. d) для пелагических видов), от относительного усилия трескового промысла (по Gislason, 1999).
Уровень Ecod, соответствующий критерию FMSY, получается выше в модели MSGVPA, в которой рост хищников зависит от количества доступной пищи. Причина этого в том, что более высокому уровню Ecod соответствует более низкая численность запаса трески, при этом увеличивается темп роста особей, поэтому несколько возрастает уровень биомассы запаса. Модель MSVPA такой "гибкостью" не обладает.
Аналогичные результаты получены для зависимости Epelag (усилие пелагического промысла, при котором достигается максимум суммарного улова сельди и шпрота) от усилия трескового промысла (рис. 7.17b). При увеличении интенсивности промысла трески ее запас уменьшается, при этом уменьшается естественная смертность пелагических рыб от хищничества М2, а промысловое усилие, обеспечивающее максимальный их вылов, следует увеличить.
Если построена поверхность улова в зависимости от двух величин промыслового усилия (пелагического и трескового промыслов), то значениям промыслового усилия по критерию F01 соответствуют точки на поверхности улова, в которых наклон поверхности равен 1/10 от ее наклона в начале координат. Обе многовидовые модели дают более высокие значения промыслового усилия по этому критерию, чем одновидовая модель VPA, и в обоих случаях усилие одного промысла возрастает при росте усилия альтернативного промысла (рис. 7.17с и 7.17d).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
