
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Далее исследуется влияние временной задержки τ – длительности времени разложение вещества, поступающего со всех трофических уровней на косную компоненту, до потребляемых первым трофическим уровнем биогенов. В этом случае первое уравнение модели (1-3) заменяется следующим:
![]() .
.
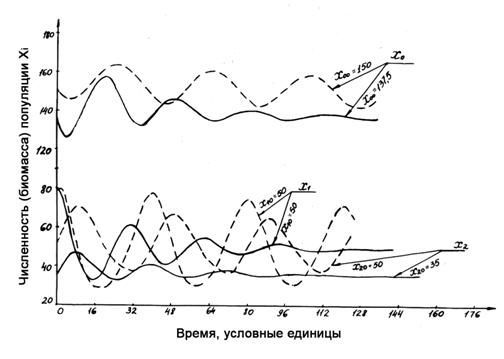
Рисунок 1.5. Сравнение варианта динамики, приведенного на рис. 1.4 (сплошные линии), с вариантом (показан пунктиром) со следующими параметрами: ε1=ε2=δ1=2*10-3, δ2=4*10-3, s1=s2=0.1, p1=p2=0.15, что соответствует ослаблению обратных связей на косную компоненту.
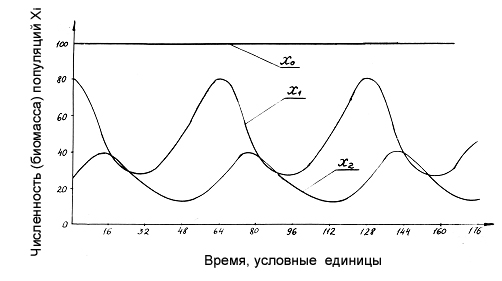
Рисунок 1.6. Динамика компонент модели (1-3), параметры которой совпадают с вариантом, приведенным пунктиром на рис.1.4, но при разрыве обратной связи со второго компонента на косный (p2=0, p1=0,2).
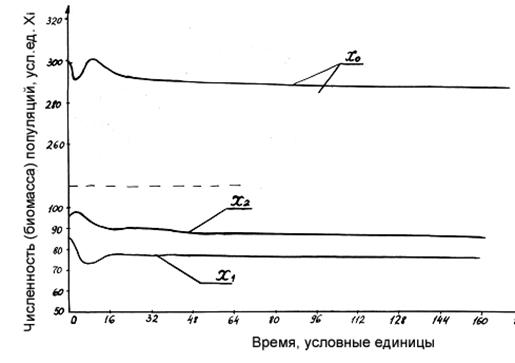
Рисунок 1.7. Динамика компонент экосистемы для n=2 при введении коэффициентов внутривидовой конкуренции.
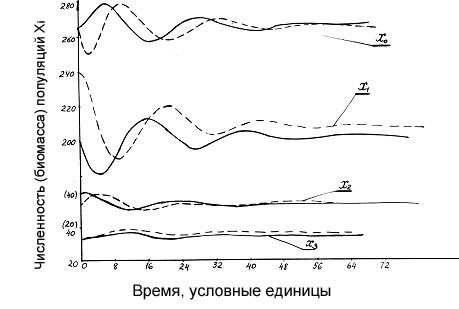
Рисунок 1.8. Динамика компонент модели (1-3) при n=3 без запаздывания (сплошные линии) и с запаздыванием (время разложения вещества τ =3) – пунктирные линии. Значения параметров модели: ε1=δ1=10-3,ε2=1,5*10-3, δ2=5*10-3, ε3=3*10-3, δ3=6*10-3, si=0,1, pi=0,2.
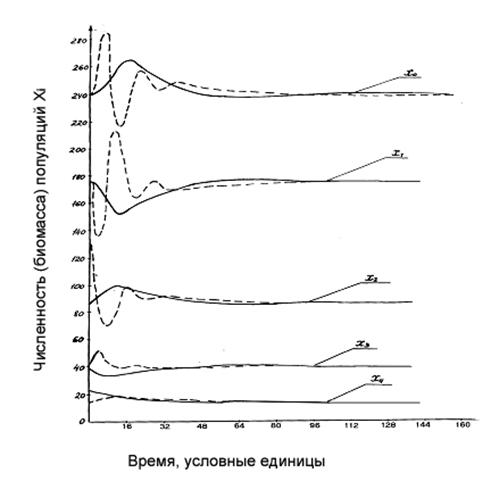
Рисунок 1.9. Динамика компонент модели (1-3) с задержкой (τ =3) при n=4.
Сплошные линии соответствуют варианту с начальным возмущением только четвертой компоненты (i=4), а пунктирные – варианту с начальным возмущением компоненты i=2. Значения параметров модели следующие: pi=0,2, si=0,1; остальные приведены в таблице:
i | 1 | 2 | 3 | 4 |
δi*103 | 1,5 | 3 | 2 | 2,5 |
εi*103 | 1,5 | 1,7 | 5 | 5 |
Расчеты при n=3 и τ=3 (рис. 1.8) показали, что при введении задержки такого типа существует устойчивая точка равновесия, при этом динамика биомассы каждого уровня представляет собой затухающие колебания с задержкой по фазе относительно аналогичного варианта расчетов при τ=0.
Следующие численные расчеты проведены для n=4 с тем же временем задержки τ=3. При начальном возмущении компоненты более высокого трофического уровня система быстрее возвращается к состоянию равновесия, чем при возмущении компоненты более низкого уровня (рис. 1.9). Этот вывод следует учитывать при эксплуатации промыслового сообщества – вмешательство (например, вылов) в нижние трофические уровни опаснее, чем в верхние уровни. К тому же выводу приходим и в следующих главах данной работы.
1.2.3. Влияние связи с косной компонентой типа «поток» на динамику популяций трофической цепи
Рассмотрим теперь случай, когда для первой компоненты экосистемы (например, для фитопланктона) лимитирующим ее развитие фактором является не количество питательных веществ в косной компоненте, а поток некоторого необходимого вещества (например, биогенов) или поток солнечной энергии (Булгакова, 1966). Схема модели приведена на рис. 1.3с. Пусть величина этого потока постоянна и равна P. Это означает, что при увеличении численности или биомассы первой компоненты (потребителя этого вещества или энергии) на единицу плотности этой компоненты будет приходиться меньшая доля притекающего вещества или энергии.
Полагаем, что прирост биомассы первого вида за малый промежуток времени Δt пропорционален его биомассе и количеству энергии или питательного вещества γ, приходящегося на единицу биомассы (или на одну особь), т. е. равен ε*x1γ. Пусть с ростом x1 потребляемое в единицу времени количество биогенов меняется согласно уравнению P(1-exp(-kx1)), как показано на рис. 1.10, тогда γ =P(1-exp(-kx1))/x1. Параметры k и ε*– константы, потому ε1’= ε*P=const.
Если остальные компоненты сообщества развиваются по тем же законам, что и в предыдущих моделях, то система уравнений модели принимает вид
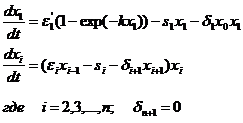 (1-8)
(1-8)
По сравнению с моделью Вольтерра (1-1) изменяется только первое слагаемое в уравнении для первого вида. Исследование модели (1-8) показало, что система с ограниченным питанием особей первого трофического уровня типа «поток» имеет единственную точку равновесия. Условие существования ее в положительной области сводится к одному неравенству xn0>0 и является одновременно необходимым и достаточным условием асимптотической устойчивости этой точки (Булгакова, 1966). Таким образом, влияние абиотического фактора описанного типа «поток» тоже играет стабилизирующую роль в динамике экосистемы.
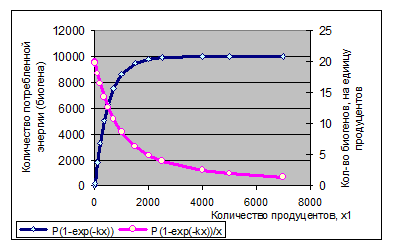
Рисунок 1.10. К модели (1-8): вид зависимости количества потребляемого биогенного вещества или солнечной энергии из потока P=const от численности или биомассы вида продуцентов x1 (левая ось ординат) и количество того же вещества, приходящееся на единицу биомассы (или на одну особь) продуцентов (правая ось ординат). Графики построены при P=10000, k=0.02.
Несколько позже появилось несколько работ с соавторами (Алексеев, 1973a, 1973б; Алексеев и Костин, 1974; Алексеев, 1980). Рассматривается в частности модель для двух трофических уровней (продуценты и консументы) при наличие косной компоненты. При этом (1973a) делает предположения, которые справедливы в случае, если xi для всех i измеряются в одних и тех же единицах, а именно – в количестве вещества, которое лимитирует развитие видов (например, фосфора или азота). Эти предположения заключаются в равенстве δ1=ε1 (все потребленное вещество косной компоненты преобразуется в биомассу первого трофического уровня), и приток вещества на косную компоненту складывается из притоков трупов каждого из видов (s1x1+s2x2) и неусвоенной видами-консументами пищи
(δ2-ε2)x1x2. В результате таких предположений получается, что изменение во времени общего суммарного лимитирующего вещества в биосистеме равно нулю, т. е. суммарное количество вещества в системе постоянно. Это условие закона сохранения вещества в биосистеме значительно упрощает исследование на устойчивость точек равновесия. В данной модели уравнение для нулевой компоненты (биогены типа «запас») принимает вид (используем наши обозначения переменных):
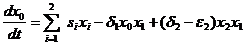 .
.
Остальные два уравнения совпадают с (1-8). Обозначим сумму количества выбранного биогенного вещества в экосистеме через Х=x1+x2+x0. Показано, что существует единственная положительная точка равновесия, которая устойчива при условии, что этого вещества в системе достаточно:
![]() .
.
Из полученного неравенства можно сделать вывод, полученный нами ранее для системы (1-8) , что с ростом δ1 –притока вещества на уровень продуцентов, или, что то же самое – при усилении связи первой компоненты с косной компонентой увеличивается устойчивость экосистемы.
Можно рассматривать и более сложную структуру косной компоненты, разделяя ее на составляющие: опад, редуценты (организмы, минерализующие органическое вещество) и биогены, – и для каждой составляющей записывать отдельное дифференциальное уравнение (Алексеев, 1973б).
Косная компонента типа «проток» исследована и (1973) в применении к бактериально-протозойным культурам. Они рассматривают модель динамики системы, состоящей из двух трофических уровней и питательной среды. Понятие «проток» означает, что в систему с постоянной скоростью втекает среда с питательными для первого уровня веществами и вытекает, унося с собой часть биомассы каждой компоненты. Авторы приходят к выводу, что такая система стабильна, т. е. существует ненулевое состояние ее равновесия, которое является либо устойчивым узлом, либо устойчивым фокусом.
1.2.4. Влияние насыщения в питании хищника на динамику системы "хищник-жертва"
В описанных выше моделях (1-3,1-5) предполагалось, что скорость потребления корма одной особью r (называемая рационом) пропорциональна количеству корма ri=δixi-1, что справедливо только в условиях голода потребителей корма. В модели же Вольтерра (1-1) предполагается, что первый вид находится в условиях изобилия корма, т. е. его рацион r1=ε1 постоянен, а последующие виды – в условиях голода.
(1955) предложил обобщенное уравнение, описывающее зависимость рациона от плотности корма, которое работает и в условиях голода и в условиях изобилия пищи, а именно ![]() , где R – максимальный рацион особи, ξ – константа, определяемая эмпирически. Решение этого дифференциального уравнения имеет вид
, где R – максимальный рацион особи, ξ – константа, определяемая эмпирически. Решение этого дифференциального уравнения имеет вид
![]() .
.
Полученная зависимость хорошо подтверждается экспериментами. Запишем модель хищник-жертва для двух видов в следующем виде:
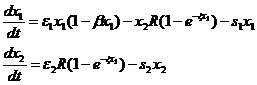 (1-9)
(1-9)
Параметр β характеризует внутривидовую конкуренцию первого вида (то есть отрицательную обратную связь – зависимость скорости роста от плотности вида), это означает, что в отсутствии хищника первый вид развивается по логистическому закону, а не по экспоненте, как предполагалось в предыдущих моделях. Формула для рациона использовалась и ранее при построении математической модели водного сообщества (Винберг и Анисимов, 1969; Виноградов и др., 1973). В данном же разделе рассматриваются лишь вопросы устойчивости модели хищник-жертва (1-9) с учетом такой зависимости (Булгакова, 1977).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
