
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Для некоторых категорий жертв были получены нулевые оценки их частного содержания в желудке ![]() (i, a;j, b;Y, Q). Их происхождение следует исследовать дополнительно, поскольку они могут отражать действительно существующую ситуацию отсутствия потребления данной категории жертвы (j, b) хищниками категории (i, a) в масштабах всего запаса хищника, а могут быть и следствием недостаточного количества проб. Естественно, в первом случае нулевые оценки следует оставлять без изменений, а во втором их следует заменять оценками, полученными с использованием процедур интерполяции и экстраполяции. К сожалению, в данном случае весьма сложно выбрать минимальное количество проб, позволяющих говорить о представительности оценок (вообще говоря, БД редко содержит гарантированно представительную информацию о питании в масштабах всего ареала хищника и жертвы с учетом всех характеристик их распределений, особенностей поведения и т. д.). В связи с этим достаточно произвольно была установлена граница представительности выборки: количество желудков хищника данного возраста, собранных в данном квартале данного года, не должно быть меньше, чем 5m, где m - общее количество возрастных групп жертвы данного вида. Например, для мойвы как жертвы в модель MSVPA включены 6 возрастных групп, потому минимальное необходимое количество желудков было принято равным 30.
(i, a;j, b;Y, Q). Их происхождение следует исследовать дополнительно, поскольку они могут отражать действительно существующую ситуацию отсутствия потребления данной категории жертвы (j, b) хищниками категории (i, a) в масштабах всего запаса хищника, а могут быть и следствием недостаточного количества проб. Естественно, в первом случае нулевые оценки следует оставлять без изменений, а во втором их следует заменять оценками, полученными с использованием процедур интерполяции и экстраполяции. К сожалению, в данном случае весьма сложно выбрать минимальное количество проб, позволяющих говорить о представительности оценок (вообще говоря, БД редко содержит гарантированно представительную информацию о питании в масштабах всего ареала хищника и жертвы с учетом всех характеристик их распределений, особенностей поведения и т. д.). В связи с этим достаточно произвольно была установлена граница представительности выборки: количество желудков хищника данного возраста, собранных в данном квартале данного года, не должно быть меньше, чем 5m, где m - общее количество возрастных групп жертвы данного вида. Например, для мойвы как жертвы в модель MSVPA включены 6 возрастных групп, потому минимальное необходимое количество желудков было принято равным 30.
6.2.4. Оценка суточных и квартальных рационов трески разных возрастов
Важным параметром модели MSVPA является средний за квартал рацион хищника R(i, a,Y, Q), который представляет собой количество пищи, потребляемой одной особью хищника вида i возраста a в течение расчетного шага (квартала Q) для года Y. При моделировании сообщества Северного моря этот параметр задавался для каждого вида хищников переменным по возрастам и кварталам, но один и тот же для всех лет.
Существует большое количество методов оценки рационов рыб, в том числе и северо-восточной арктической трески (Temming, 1986; Краснопер, 1988; Орлова, 1989; Bogstad and Mehl, 1990; Dolgov et al.,1992). В основе большинства моделей, применяемых для оценки рациона, лежит уравнение изменения массы содержимого желудка в процессе переваривания пищи, а именно:
dW(t)/dt =R(t) –fe (t) , (6-7)
где W(t) – масса пищевого комка, и скорость ее изменения равна разности между скоростью потребления пищи R(t) и скоростью ее эвакуации fe(t). При оценке среднего рациона за некоторый интервал времени предполагают, что в течение этого времени выполняется условие равновесия – т. е. что эти две скорости равны. Тогда величина рациона определяется через скорость эвакуации пищи, и различие между моделями сводится главным образом к разным предположениям о механизме переваривания пищи, т. е. к разным видам функции fe(t) (Краснопер, 1988).
Чтобы оценить скорость эвакуации пищи трески, норвежские ученые Х. дос-Сантос и М. Джоблинг (dos Santos and Jobling, 1992) провели серию экспериментов по питанию трески и определили эмпирические зависимости скорости эвакуации от различных факторов – от температуры воды T, массы хищника WP и от исходной массы проглоченной пищи Wo. Пусть H – период полу-переваривания (время, за которое переваривается половина исходной массы пищи), который выражается в виде эмпирической функции этих трех переменных параметров и двух констант (c=0,1085 и b=0.5385):
H=Hjexp(-cT)(W0j/ WP)b (6-8)
где Hj – константы полу-переваривания (выраженные в часах), различные для разных видов жертв j. Дос Сантос (dos Santos, 1990) получил оценки этого параметра для четырех видов жертв: эвфаузиидов, сельди, мойвы и креветки.
Предположив, что процесс переваривания каждого вида жертв происходит с постоянной скоростью (принята экспоненциальную модель переваривания), Б. Богстад и С. Мель (Bogstad and Mehl, 1992) получили следующие формулы для расчета суточных рационов:
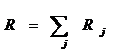 – общий суточный рацион равен сумме частных, где частный суточный рацион (потребленная за сутки биомасса жертвы j-го вида) равен
– общий суточный рацион равен сумме частных, где частный суточный рацион (потребленная за сутки биомасса жертвы j-го вида) равен
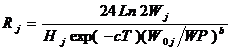 (6-9)
(6-9)
Вслед за Б. Богстадом и С. Мелем принимаем следующие значения констант Hi: для трески как жертвы и для сельди – 452, для креветки – 533, для мойвы – 283, для другой пищи трески-хищника в возрасте одного и двух лет – 205 (как для эвфаузиид), а для более старшей трески, у которой преобладает рыбная пища – 452.
Преимущество данной модели состоит в том, что она позволяет учитывать различную скорость переваривания разных видов жертв, при этом константы Hi заранее известны. При расчете квартального рациона для каждого года и каждого возраста трески в формулу (6-9) подставляется средний вес пищевого комка, средние доли в нем (по массе) каждого вида жертв и другой пищи, средний вес тела хищника (все эти значения рассчитаны по БД) и средняя температура в районе питания хищника.
Поскольку в БД нет данных о температуре на станции, где поймана треска, для расчетов рациона использовались данные по донной температуре, усредненные по всему району для данного интервала времени. Таким образом, для всех возрастных групп трески в фиксированном интервале времени в расчетах использовалась одна и та же температура, что вносило некоторую погрешность в оценки рационов.
Квартальный рацион определяется умножением суточного рациона на среднее количество дней в квартале (90,25). Полученные значения квартальных рационов трески в возрасте 1-10 лет по кварталам от 1984 г. и по 1991 г. приведены в нашей статье (Булгакова и др., 1995, табл. 3). Изменчивость рациона по кварталам (сезонная изменчивость) и по годам сильно связана с изменчивостью кормовой базы. На рис. 6.14 показана динамика относительных суточных рационов (отношение массы пищи, съеденной за сутки одной особью, к массе тела хищника). Видно, что существенна не только сезонная, но и межгодовая изменчивость рационов трески. Это свойство бореальной экосистемы заставило нас отказаться от предыдущей версии модели, в которой было заложено постоянство рационов по годам. В рамках международного гранта1[8]Х. Гисласон (Дания) модифицировал модель MSVPA специально для баренцевоморской ЭС, позволив представлять рацион трески в виде функции трех переменных R(a, Y,Q).
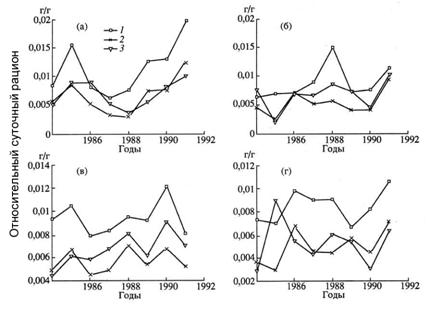
Рисунок 6.14. Годовая динамика относительных суточных рационов (г/г) для трех возрастных групп трески: три года (1), пять (2) и семь (3) лет для интервала 1984-1991 гг. Рисунки а-г соответствуют кварталам I-IV (Булгакова и др., 1995).
Всесторонний анализ динамики рационов трески проведен в наших работах (Булгакова и др., 1995; Bulgakova et al., 1995a). Из четырех кварталов хуже всего в БД представлен четвертый. Доля пустых желудков самая большая в I и II кварталах. Анализ частных рационов показал, например, что в первом квартале изменение доли мойвы по массе в рационе происходит синхронно с изменением запаса мойвы, во II – IV кварталах увеличивается роль креветки и резко возрастает роль "другой пищи", особенно в годы депрессии запаса мойвы (в 1988 г. доля другой пищи достигала 95% рациона).
Полученные нами оценки относительного суточного рациона лежат в интервале от 0,4% до 2,0%. Эти оценки близки к оценкам, полученным с соавторами (Dolgov et al., 1992) – 0,54-2,2%. В ряде работ, где применялись другие методы оценки рационов или расчет велся для отдельного района или определенного периода откорма трески, оценки суточного рациона иногда превышали 4% (Тарвердиева и Ярагина, 1989; Orlova, 1992).
При определении рациона по модели (6-9) используется несколько параметров, оценка каждого из них несет в себе ошибку, дополнительная ошибка вносится также при усреднении. и (1999) при подготовке входных данных по квартальным рационам для каждого временного интервала (год-квартал) предложили сглаживать полученные оценки логистической функцией от возраста трески, которая зависит от трех параметров b0, b1 и b2:
R (a, Y, Q) = b0 /(1+b1EXP(b2a)), (6-10)
Параметры b0, b1 и b2 определяются в процессе аппроксимации данных, рассчитанных по формулам (6-9) для выбранного года и квартала (рис. 6.15). Такая аппроксимация может служить также инструментом для экстраполяции величины рациона на старшие возрастные группы, для которых, как правило, мало данных по составу желудков.
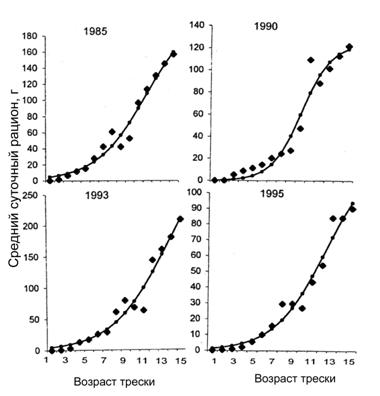
Рисунок 6.15. Суточные рационы трески (в г) в зависимости от возраста и их аппроксимация логистической зависимостью в первом квартале для четырех лет (по Коржеву и Долгову, 1999)
и (1999) расширили модель сообщества Баренцева моря, включив в нее второго хищника–пикшу, а также двух "внешних хищников" – гренландского тюленя и малого полосатика. Поскольку для пикши эксперименты по определению скоростей переваривания разных видов пищи не проводились, авторы рассчитывали рационы пикши по методу П. Бромли (Bromley, 1991).
6.2.5. Оценка коэффициентов “остаточной” смертности
Оценка коэффициента “остаточной” естественной смертности является чрезвычайно сложной задачей. Едва ли существует достаточно надежный способ точной ее оценки. Эта составляющая естественной смертности (М1) для младших возрастных групп действует в модели одновременно со смертностью от хищничества (М2), а последняя тоже заранее неизвестна и оценивается внутри процедуры расчетов MSVPA. По этой причине используемые в моделях значения параметра M1 являются по сути не более чем в той или иной мере разумной догадкой, сделанной на основе анализа дополнительной информации.
Новый способ подбора параметра M1 предложено использовать (Многовидовой анализ.., 2001) для таких популяций, у которых хищничество или каннибализм влияют только на младшие возрастные группы (в нашем случае это треска и сельдь). Для возрастов старше трех лет можно считать параметр M1 равным принятому постоянному мгновенному коэффициенту естественной смертности (в ИКЕС для трески этот коэффициент принят равным 0,2 [1/год]), и распределить его равномерно по кварталам, то есть M1=0,05 [1/квартал]. Этот способ заключается в одновременной настройке параметра M1 с терминальными коэффициентами промысловой смертности многовидовой модели с помощью одновидовой сепарабельной модели ISVPA (см. раздел 6.3). Для трески моложе трех лет оценки M1 получены из материалов, используемых и др. (1998) при построении модели поколения трески (табл. 6.9).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
