
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
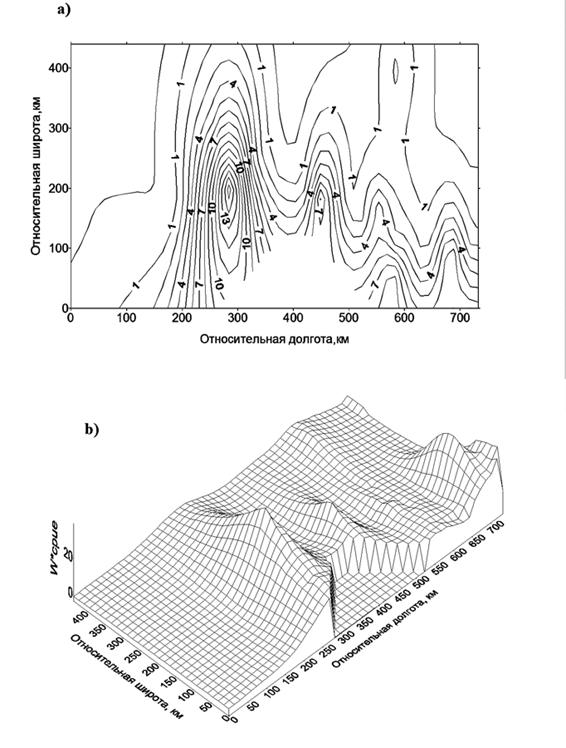
Рисунок 6.7. Изолинии массы трехлетней мойвы в желудках трехлетней трески (г) (а) и поверхность пространственного распределения (b) произведения массы трехлетней мойвы в желудках на количество особей трески в возрасте 3 лет, пойманной за час траления (cpue). Все относится к первому кварталу 1990г. Начало координат, как на рис.6.2.
Для отдельной возрастной группы может оказаться недостаточно данных для проведения такого анализа. В случаях, если имеющаяся в БД информация не позволяет провести пространственный анализ данных по питанию (например, количество проб слишком мало или они взяты в слишком узкой части ареала), описанный выше подход не имеет преимуществ по сравнению с традиционным арифметическим усреднением со взвешиванием по cpue. В некоторых случаях (например, когда радиус поиска в рамках процедуры кригинга мал), результаты расчетов с учетом пространственных факторов совпадают с результатами арифметического усреднения со взвешиванием по cpue.
6.2.3.2. Применение специальной методологии для заполнения пробелов в базе данных по питанию хищников
Как правило, достаточно представительная информация по питанию трески содержится в БД лишь для возрастных групп не старше 8-10 лет. Информация по питанию более старшей трески встречается лишь эпизодически: она может быть представлена единичными желудками, часто пустыми или содержащими экстремально большое количество жертв какой-либо одной категории. Высокая изменчивость состава пищи и темпов питания между отдельными особями требует достаточно большого размера проб. Непосредственное использование в качестве входных данных для МSVPA оценок, полученных по непредставительным данным, может привести к ошибочным результатам.
Заранее определить вид и параметры возрастной зависимости общей массы и состава пищи в желудках достаточно сложно. Тем не менее, представляется разумным основываться на следующих предположениях (Bulgakova, Vasilyev, Daan, 2001):
· общая масса содержимого желудков ![]() tot(a) должна увеличиваться с возрастом, а поскольку в старших возрастах наблюдается снижение темпа роста массы особей, скорость увеличения массы содержимого желудков также должна уменьшаться с возрастом, возможно, по асимптотическому закону; но параметры этой зависимости могут отличаться от параметров функции роста массы тела, поскольку изменения энергетического обмена в старших возрастных группах могут привести к более быстрому выходу кривой на плато;
tot(a) должна увеличиваться с возрастом, а поскольку в старших возрастах наблюдается снижение темпа роста массы особей, скорость увеличения массы содержимого желудков также должна уменьшаться с возрастом, возможно, по асимптотическому закону; но параметры этой зависимости могут отличаться от параметров функции роста массы тела, поскольку изменения энергетического обмена в старших возрастных группах могут привести к более быстрому выходу кривой на плато;
· средняя масса пищи в желудках зависит от условий питания, что может приводить к межгодовым и межквартальным различиям в параметрах;
· формализация возрастной зависимости частного содержимого желудков является ещё более сложной процедурой, поскольку и пищевые предпочтения хищника с возрастом могут меняться.
На основе этих предположений была предпринята попытка описать зависимость общей массы пищи в желудке от возраста логистической кривой, отдельно для каждого квартала каждого года. К сожалению, в большинстве случаев не удалось оценить параметры с приемлемой надежностью, поскольку имеющиеся данные описывают, как правило, только начальную часть кривой и не дают достаточной информации об ее асимптотических свойствах. Более устойчивые оценки были получены при обработке двумерных массивов, составленных из оценок ![]() tot(a;Y) как функции двух переменных (возраста хищника и года) отдельно для каждого квартала. Исходное предположение для применения такой процедуры состояло в том, что дефицит информации для отдельных возрастных групп в каком-либо году может быть в определенной степени компенсирован информацией за смежные годы. Таким образом, задача сводится к аппроксимации двумерного массива данных некоторой поверхностью, достаточно гибкой для отражения возрастных и межгодовых тенденций в изменениях средней массы пищи в желудке, но в то же время достаточно устойчивой к воздействию случайных выбросов в оценках.
tot(a;Y) как функции двух переменных (возраста хищника и года) отдельно для каждого квартала. Исходное предположение для применения такой процедуры состояло в том, что дефицит информации для отдельных возрастных групп в каком-либо году может быть в определенной степени компенсирован информацией за смежные годы. Таким образом, задача сводится к аппроксимации двумерного массива данных некоторой поверхностью, достаточно гибкой для отражения возрастных и межгодовых тенденций в изменениях средней массы пищи в желудке, но в то же время достаточно устойчивой к воздействию случайных выбросов в оценках.
Имеющиеся в БД данные для такого анализа представлены на рис. 6.8. Как видно из рисунка, несмотря на весьма большое общее количество собранных проб, некоторые из возрастных групп хищника в БД представлены недостаточно. Хуже всего представлены старшие возрастные группы, а в некоторые годы полностью отсутствуют данные для возрастных групп 1 и 2.
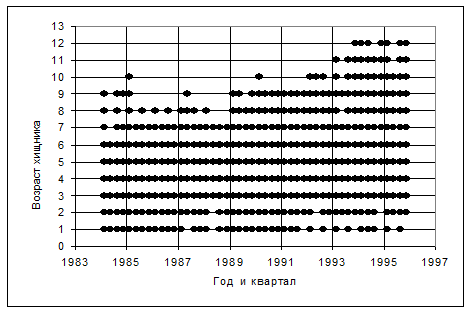
Рисунок 6.8. Наличие представительных данных по содержимому желудков (количество желудков не менее 5) по возрастам хищника (трески) и кварталам и годам для 1984-1995 гг.
Ранее, как правило, все имеющиеся в БД данные использовались в качестве входной информации для модели MSVPA, даже если информация была представлена лишь одним желудком. Если же для некоторых возрастных групп хищника информация отсутствовала вовсе, она достаточно произвольно заменялась данными по соседним возрастным группам того же квартала или же данными для этой возрастной группы за какой-либо соседний год. Особенно необоснованным было восполнение отсутствующих данных для старших возрастов: как правило, на более старшие группы переносились данные, имеющиеся для самой старшей репрезентативной возрастной группы. Это могло приводить к существенным смещениям оценок количества потребленных хищником жертв, а значит и оценок запасов. Следует отметить, что для проведения описанной ниже процедуры экстраполяции в качестве входных данных следует брать только такие точки на плоскости (a, Y) {возраст хищника; год}, которые обеспечены достаточным количеством собранных желудков. В данном случае мы брали только такие «точки», для которых имелось более 5 желудков. Именно такие точки показаны кружками (рис. 6.8).
Работа с вариограммами, построенными для массивов данных ![]() tot(a;Y) для выбранного квартала в направлении года и возраста, показала, что удобно заменить сами величины их логарифмами, в этом случае легче выбрать модель вариограммы. Анализ вариограмм (рис. 6.9) показал, что межгодовая изменчивость оценок общей массы пищи в желудках существенно ниже, чем межвозрастная. Это в частности подтверждает наше предположение, что для заполнения пробелов в данных следует использовать статистически обоснованную методологию экстраполяции, а не присваивать одно и то же значение массы пищи старшим возрастным группам, для которых нет фактических данных.
tot(a;Y) для выбранного квартала в направлении года и возраста, показала, что удобно заменить сами величины их логарифмами, в этом случае легче выбрать модель вариограммы. Анализ вариограмм (рис. 6.9) показал, что межгодовая изменчивость оценок общей массы пищи в желудках существенно ниже, чем межвозрастная. Это в частности подтверждает наше предположение, что для заполнения пробелов в данных следует использовать статистически обоснованную методологию экстраполяции, а не присваивать одно и то же значение массы пищи старшим возрастным группам, для которых нет фактических данных.
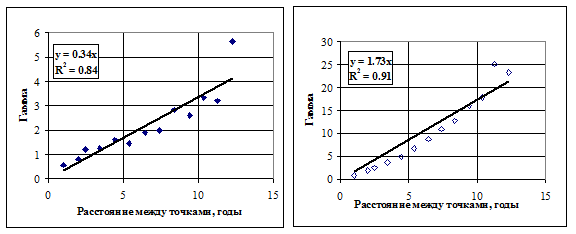
Рисунок 6.9. Вариограммы Ln[W(a, Y)] для первого квартала, построенные в двух направлениях: года (слева) и возраста трески-хищника (справа). Прямые линии соответствуют линейным трендам.
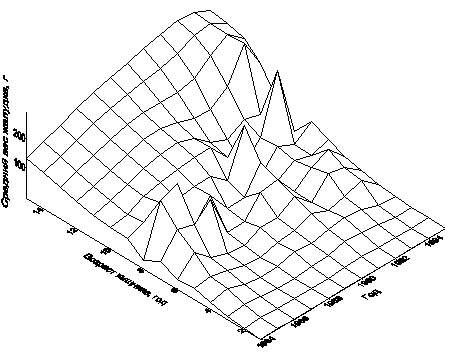
Рисунок 6.10. Построенная с помощью кригинга поверхность Wtot(a, Y)
в граммах как функция возраста хищника и года для первого квартала
Поверхности зависимости общей массы содержимого желудка трески от двух переменных, года и возраста хищника, получены с помощью кригинга для каждого из четырех кварталов (Булгакова, Васильев, 1998). Для первого квартала такая поверхность приведена на рис. 6.10. Сечение этой поверхности плоскостью при фиксированном годе Y=const дает зависимость массы содержимого желудка трески от её возраста для этого года. Для четырех произвольно выбранных лет представлены такие сечения (рис. 6.11).
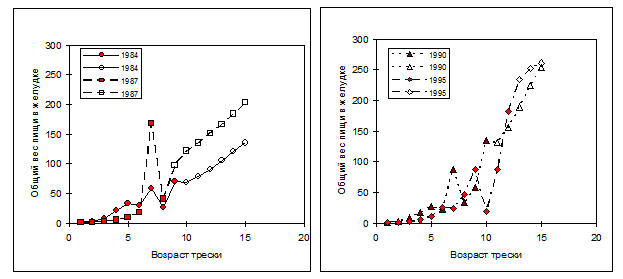
Рисунок 6.11. Наблюдаемые (закрашенные значки) и экстраполированные с помощью кригинга (незакрашенные значки) значения общего содержимого желудков трески, в г, в зависимости от ее возраста для четырех лет: 1984 и 1987 (слева) и 1990 и 1996 (справа).
Следует отметить, что при проведении кригинга мы выбирали такие опции, которые с одной стороны, соответствовали полученной модели вариограммы, а с другой – не вносили искажений в те точки, для которых имелось достаточное количество желудков.
Задача статистически обоснованного заполнения пробелов в массивах информации возникает также при подготовке входных данных по частному составу пищи ![]() (i, a;j, b;Y, Q). Для ее решения для каждого сочетания вида хищника и вида жертвы и каждого интервала времени создаются двумерные массивы {Wst(a, b)i, j,Y, Q} и исследуются построенные для них вариограммы. Затем c помощью кригинга созданные массивы аппроксимируют функцией двух переменных (возраст хищника и возраст жертвы). Логика применения такой процедуры заключается в стремлении компенсировать дефицит информации по потреблению рассматриваемой категории жертвы (j, b) информацией о ее потреблении другими возрастными группами хищника, а также данными по потреблению рассматриваемой возрастной группой хищника жертв других возрастов того же вида в данном квартале рассматриваемого года.
(i, a;j, b;Y, Q). Для ее решения для каждого сочетания вида хищника и вида жертвы и каждого интервала времени создаются двумерные массивы {Wst(a, b)i, j,Y, Q} и исследуются построенные для них вариограммы. Затем c помощью кригинга созданные массивы аппроксимируют функцией двух переменных (возраст хищника и возраст жертвы). Логика применения такой процедуры заключается в стремлении компенсировать дефицит информации по потреблению рассматриваемой категории жертвы (j, b) информацией о ее потреблении другими возрастными группами хищника, а также данными по потреблению рассматриваемой возрастной группой хищника жертв других возрастов того же вида в данном квартале рассматриваемого года.
Результат проведения кригинга показан на примере содержимого мойвы в желудках хищника в первом квартале 1990 г. (рис.6.12). За этот квартал одна особь семилетней трески потребила более 40 г четырехлетней мойвы – существенно больше, чем в других возрастах.
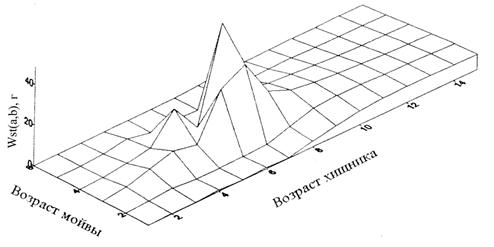
Рисунок 6.12. Полученная с помощью кригинга поверхность частного содержимого мойвы в желудках трески как функция возраста трески и возраста мойвы для первого квартала 1990 г.
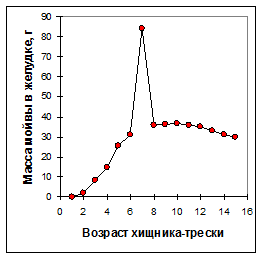
Рисунок 6.13. Суммарная масса мойвы (по всем ее возрастам) в среднем желудке хищника-трески в зависимости от возраста трески (I квартал 1990 г.).
По результатам кригинга построена зависимость суммарного содержания мойвы всех возрастов в желудках хищника в зависимости от возраста трески для первого квартала 1990 года (рис.6.13). Очень высокое значение массы мойвы Wst в желудках семилетней трески похоже на выброс, но оно обеспечено большим количеством исследованных желудков трески этого возраста (460). Остальные же значения укладываются в весьма гладкую кривую, и экстраполяция выглядит правдоподобной.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
