
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Стационарная точка модели (1-9) определяется как
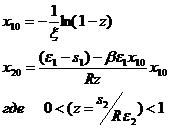 (1-10)
(1-10)
Эта точка лежит в положительной области при выполнении условия
![]()
При β=0 это условие сводится к неравенству R>s2/ε2, которое эквивалентно условию z<1.
Исследуем на устойчивость модель (1-10). В системе уравнений (1-9) делаем замену переменных xi=yi+xi0, затем проводим линеаризацию полученных уравнений, отбрасывая члены порядка выше первого и используя приближенное представление экспоненты возле начала координат. Учитываем при этом соотношение exp(-ξx10)=1-z . Полученная линеаризованная система уравнений (1-11) описывает динамику сообщества вблизи точки равновесия.
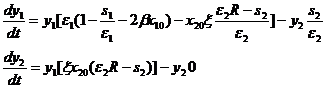 (1-11)
(1-11)
Характеристический определитель системы (1-11) имеет вид
![]() , (1-12)
, (1-12)
где
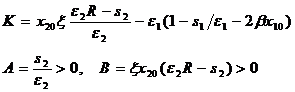 (1-13)
(1-13)
Коэффициенты A и B положительны при условии, что точка равновесия (1-10) лежит в положительной области. Определители Гурвица (1-12) для характеристического уравнения положительны при условии K>0, поскольку определитель (1-12) является частным случаем определителя (13) из нашей работы (Булгакова, 1966). Это означает, что при выполнении условия K>0 система (1-9) имеет асимптотически устойчивую точку равновесия. Рассмотрим подробнее условие K>0. Подставив выражение для x20 из (1-10) в первое слагаемое неравенства, после несложных преобразований получим
![]() , где
, где ![]() .
.
Из второго уравнения системы (1-10) ясно, что f-βx10>0, и после деления на это выражение получаем неравенство
![]() (1-14)
(1-14)
Обозначим левую часть неравенства (1-14) через L(z), правую – через F(z). Исследование показывает, что при условии 0<z<1 функция L(z) уменьшается на этом интервале от 1 до нуля (рис. 1.11). Функция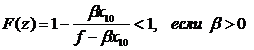 , поскольку согласно условию (1-10) выполняется неравенство (f-βx10)>0.
, поскольку согласно условию (1-10) выполняется неравенство (f-βx10)>0.
Условие устойчивости модели «L(z)>F(z)» начинает выполняться при увеличении значений параметра β. Функции F(z) построены при следующих значениях параметров: f=0,7; ε1=0,4; ξ=0,01.
При β =0 (отсутствие внутривидовой конкуренции вида жертв) F(z)=1, а функция L(z)<1 на всем интервале значений z, т. е. условие (1-14) не выполняется, значит, что модель (1-9) неустойчива. Это можно объяснить тем, что существование максимального рациона хищника R не позволяет хищнику регулировать численность жертв так, как при неограниченном рационе в модели Вольтерра. Если в модели Вольтерра сообщество находилось на границе устойчивости, то теперь оно становится неустойчивым.
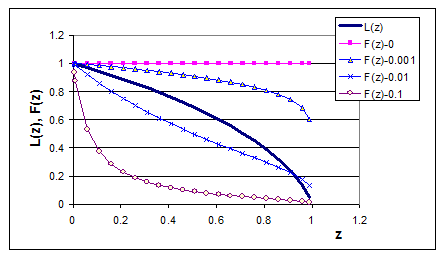
Рисунок 1.11. Иллюстрация к исследованию устойчивости модели «хищник-жертва» с учетом насыщения питания хищника. L(z) – левая часть неравенства (1-14) как функция обобщенного параметра z, семейство функций F(z) – правая часть того же неравенства при различных значениях параметра внутривидовой конкуренции вида жертв β, указанных в легенде.
Функция F(z) зависит от параметра β, и легко видеть, что производная этой функции по β отрицательна, то есть F(z) уменьшается при увеличении коэффициента β. Начиная с некоторого значения параметра β (см. рис. (1.11)), начинает выполняться условие устойчивости системы (1-14).
Таким образом, условие устойчивости стационарной точки модели (1-9), в которой взаимоотношения видов описываются более близкой к реальности зависимостью, чем в модели Вольтерра, существует только при учете в модели внутривидовой конкуренции первого вида.
с соавторами (1972) предложили модель хищник-жертва, в которой зависимость рациона от численности и хищников и жертв записывается в более общем виде, а именно, в виде функции от отношения численностей видов r(x1/x2) , которая должна удовлетворять условиям:
r(0)=0; r(∞)=R=const; 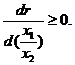
Этим условиям удовлетворяет, например, предложенная (1955, стр.129) функция
![]() . (1-15)
. (1-15)
Включение в зависимость рациона от количества корма еще одной переменной – численности хищников (переменной x2) как будто позволяет учесть внутривидовую конкуренцию хищников, но такое усложнение приводит к отсутствию в модели состояния равновесия. Рассмотрим, например, модель Гинзбурга и др.(1971) для двух видов при условии существования зависимости (1-15):
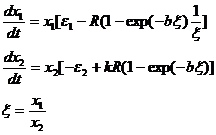 (1-16)
(1-16)
Точки равновесия модели (1-16) существуют и расположены на луче x1=ξ0x2, если два независимых алгебраических уравнения (которые получаются при условии, что правые части системы (1-16) равны нулю) имеют один и тот же корень ξ0, тогда . Подставляя это выражение в любое из уравнений (1-16), получим равенство
![]() .
.
Несомненно, что математически такое соотношение между параметрами популяций может выполняться, но биологически это условие слишком жесткое, поскольку требует равенства функций от нескольких параметров моделируемой системы. Такое соотношение будет нарушаться при любом случайном отклонении значений любого из входящих в него параметров.
Отметим, что запись условий существования равновесия системы в виде равенств функций от параметров встречается и в других работах (например, в работе с соавторами (1966)) и практически означает, что равновесное состояние в изучаемой модели отсутствует. Это означает, что исследуемая модель плохо соответствует оригиналу.
(1973) и (1973в) в модели «хищник-жертва» используют другую зависимость рациона от количества корма, а именно:
![]() , где α и b – постоянные параметры.
, где α и b – постоянные параметры.
Графически эта зависимость очень близка к зависимости , используемой нами в модели (1-9). Показано (Базыкин, 1973), что в модели «хищник-жертва» и при этой зависимости рациона система неустойчива, если не учитывать внутривидовую конкуренцию, а при учете внутривидовой конкуренции система может иметь одну, а иногда и две устойчивые точки равновесия.
1.2.5. Обобщенный подход к моделированию трофической цепи
Во всех перечисленных выше математических моделях правые части дифференциальных уравнений описываются конкретными функциями общего типа. Существует также ряд работ, в которых правые части уравнений явно не заданы, но перечисляются свойства этих функций, обеспечивающие разные типы взаимодействия видов. Из этих качественных предпосылок авторы получают качественные оценки решения.
Впервые такие модели рассматривал (Kolmogoroff, 1936; Колмогоров, 1972), который представил модель «хищник-жертва» в виде:
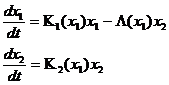 (1-17)
(1-17)
Относительно функций К1, К2 и Λ были сделаны следующие гипотезы:
1) эти функции непрерывны и непрерывно дифференцируемы;
2) dK1/dx1<0 (т. е. ограничены ресурсы для существования первого вида), при этом K1(0)>0>K1(∞)>-∞;
3) Λ>0 при x1>0;
4) dK2/dx1>0 (с ростом числа жертв увеличивается скорость размножения хищников): K2(0)<0<K2(∞).
Отметим, что модель Вольтерра (1-1) не удовлетворяет перечисленным свойствам, поскольку в ней K1=ε1=const.
На основании этих гипотез получены условия устойчивости стационарных точек, расположенных в положительном квадранте.
Качественно исследуется возможность существования периодических решений системы двух уравнений следующего типа (Uts and Waltman, 1963):
![]() (1-18).
(1-18).
Авторы доказывают следующую теорему:
если для функций F и G вторые производные существуют и непрерывны, если точка (a, b) является ненулевой точкой равновесия и частные производные в этой точке ![]() не равны нулю, и если выполняется неравенство
не равны нулю, и если выполняется неравенство 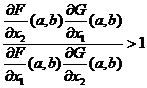 , то существует такая окрестность точки
, то существует такая окрестность точки 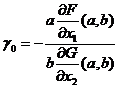 , что для любого значения параметра γ из этой окрестности существует периодическое решение системы (1-18).
, что для любого значения параметра γ из этой окрестности существует периодическое решение системы (1-18).
Система (1-18) представляет собой общий вид математической модели двух видов. Покажем, что вышеприведенная теорема формулирует достаточные, но не необходимые условия периодичности решения. Например, модель Вольтерра (1-1) для n=2 имеет периодическое решение, но не удовлетворяет условиям теоремы, поскольку функции F и G в модели Вольтерра являются функциями одного переменного, и ![]()
Опираясь на работу (Kolmogoroff, 1936), А. Рисайо и И. Ричардсон (Rescigno and Richardson, 1967) рассматривают математическую модель двух взаимодействующих видов

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
