
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
1.3. Исследование моделей конкурирующих видов
Термин «конкуренция» имеет множество трактовок, в настоящей работе этот термин употребляется в смысле А. Мильна (1964): «Конкуренция – это стремление двух (или более) животных обладать одним и тем же определенным предметом или же захватить часть, необходимую каждому, из запаса, недостаточного для удовлетворения потребностей обоих (или всех) животных». Под предметом, за который идет конкуренция, можно понимать либо количество потребляемого вещества (пищи), либо пространство. Если имеется в виду пища, то конкурирующие особи или популяции должны относиться к одному и тому же трофическому уровню.
Иногда исследователи считают виды конкурентами, если у них совпадают или очень близки спектры питания, т. е. если они питаются одной и той же пищей. Как видно из приведенного определения, для существования конкуренции этого недостаточно, необходимо, чтобы количество пищи ограничивало развитие видов.
Рассмотрим несколько математических моделей конкуренции. В. Вольтерра (Volterra, 1931; Volterra and d’Ancona, 1935) строил модель конкурирующих за пищу видов, основываясь на следующих предположениях:
а) если пища находится в достаточном количестве, то коэффициенты размножения видов εi (i=1,2,…) положительны и постоянны;
б) если количество съедаемой пищи влияет на развитие i-го вида, то его коэффициент размножения уменьшается на величину γiF(x1,x2,…xn), где γi- положительная константа, а F(x1,x2,…xn) – количество пищи, съедаемое в единицу времени всеми видами. При этом F(x1,x2,…xn)≥0, причем равенство достигается, только если для всех i выполняется равенство xi=0; limF(x1,x2,…xn)=∞ , если хотя бы для одного вида xi стремится к бесконечности.
Система уравнений этой модели конкуренции n видов имеет вид
(1-25)
Поскольку во все уравнения (1-25) входит одна и та же функция F, для двух любых видов этого сообщества с индексами r и s можно написать соотношение:
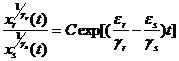 , (1-26)
, (1-26)
где C – константа, зависящая от начального состояния этих двух видов, равная 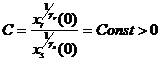 . Отсюда получаем, что при
. Отсюда получаем, что при 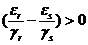 левая часть равенства (1-26) стремится к бесконечности, а поскольку численности всех видов ограничены, то вид с индексом s со временем вымирает. Составив такие соотношения для каждой пары видов, Вольтерра (Volterra, 1931) показал, что из n видов выживает один вид, для которого отношение
левая часть равенства (1-26) стремится к бесконечности, а поскольку численности всех видов ограничены, то вид с индексом s со временем вымирает. Составив такие соотношения для каждой пары видов, Вольтерра (Volterra, 1931) показал, что из n видов выживает один вид, для которого отношение ![]() наибольшее. Два вида могут существовать бесконечно долго, только если выполняется равенство
наибольшее. Два вида могут существовать бесконечно долго, только если выполняется равенство ![]() , что маловероятно. Таким образом, в системе конкурирующих видов, описанной моделью Вольтерра, все виды вымирают, кроме одного.
, что маловероятно. Таким образом, в системе конкурирующих видов, описанной моделью Вольтерра, все виды вымирают, кроме одного.
(1933, 1934а, 1934б, 1935) подробно изучал процесс конкуренции видов и в лабораторных экспериментах, и с помощью математической модели, которая является несколько более общей, чем модель Вольтерра.
Для каждого вида вводится параметр Ki , который называется емкостью среды и имеет смысл максимальной численности i-го вида в условиях отсутствия конкуренции (Гаузе, 1934a), то есть Ki особей может быть обеспечено ресурсом, лимитирующим развитие данного вида. Согласно Гаузе, скорость роста численности каждого вида зависит от потенциально возможного ее прироста и от неиспользованных возможностей для развития этого вида. В отсутствии конкуренции скорость увеличения численности запаса, развивающегося в ограниченной среде, равна ε(1-x/K), а численность популяция меняется во времени согласно логистическому закону.
При наличии вида-конкурента неиспользованная возможность развития (или неиспользованный ресурс) первого вида выразится в виде
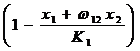 ,
,
где ![]() – коэффициент конкуренции. Смысл этого коэффициента в том, что x2 особей второго вида потребляют столько же лимитирующего ресурса, сколько ( x2) особей первого вида. Тогда систему уравнений модели конкуренции двух видов можно записать в следующем виде:
– коэффициент конкуренции. Смысл этого коэффициента в том, что x2 особей второго вида потребляют столько же лимитирующего ресурса, сколько ( x2) особей первого вида. Тогда систему уравнений модели конкуренции двух видов можно записать в следующем виде:
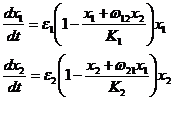 (1-27).
(1-27).
Эту систему уравнений исследовал методом изоклин (изоклины – это линии равного наклона на плоскости, где по осям координат отложены численности видов). На плоскости {x1,x2} уравнение изоклин записывается в виде dx1/dx2=const, получается оно при делении одного из уравнений (1-27) на другое. Ограничимся уравнениями горизонтальной и вертикальной изоклин (dx1/dx2 = 0 и dx2/dx1 = 0 соответственно), в данном случае они имеют вид прямых линий:
![]() ;
; ![]() . (1-28)
. (1-28)
В зависимости от разного расположения этих линий возможны четыре разные ситуации (рис. 1.14).
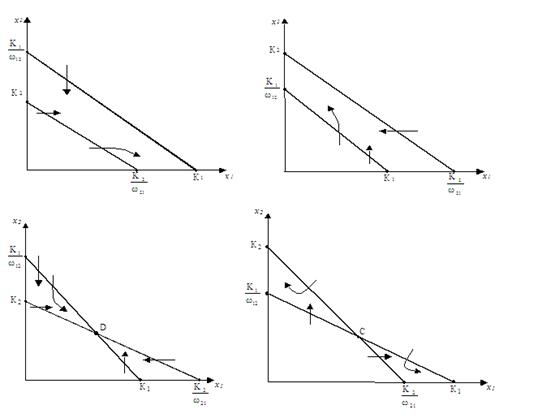
Рисунок 1.14. Четыре возможных случая расположения изоклин двух конкурирующих видов согласно модели . Стрелками показаны направления перемещения точки по интегральной кривой при пересечении изоклины.
1. ![]() – выживает только первый вид.
– выживает только первый вид.
2. ![]() – выживает только второй вид.
– выживает только второй вид.
3. ![]() – в положительной области есть точка пересечения изоклин, которая является точкой равновесия (dN1/dt = dN2/dt = 0), эта точка устойчива и является узлом, ее координаты равны
– в положительной области есть точка пересечения изоклин, которая является точкой равновесия (dN1/dt = dN2/dt = 0), эта точка устойчива и является узлом, ее координаты равны
![]()
4. ![]() . Точка пересечения изоклин существует, но неустойчива. Один из видов вымирает, другой выживает, а какой именно выживает, зависит от начальных численностей видов (Булгакова, 1970).
. Точка пересечения изоклин существует, но неустойчива. Один из видов вымирает, другой выживает, а какой именно выживает, зависит от начальных численностей видов (Булгакова, 1970).
В случаях 1 и 2 изоклины не пересекаются в положительной области значений x1 и x2. В случае 3 экологические ниши1[2]двух видов пересекаются, но не совпадают, каждый из них имеет часть ниши, в которой условия для этого вида более благоприятны, чем для его конкурента (Макфедьен, 1965). Если экологические ниши видов не перекрываются, ω12=ω21=0, т. е. конкуренции нет. Если же ниши совпадают полностью, то ω12ω21=1. Если ω12=ω21=1, то либо эти виды идентичны, либо это две группы особей одного и того же вида. Тогда и K1=K2 и x1(t)=x2(t)x1(0)/x2(0), т. е. численности этих видов меняются по одному и тому же закону и относятся как начальные их численности.
Покажем, что модель конкуренции Вольтерра является частным случаем модели Гаузе. Для этого перепишем систему уравнений Гаузе (1-27) в виде:
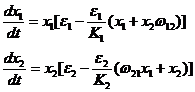
Обозначим ![]() . Поскольку в модели Вольтерра F1=F2, то для всех значений xi должно выполняться равенство
. Поскольку в модели Вольтерра F1=F2, то для всех значений xi должно выполняться равенство 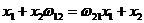 , что означает равенство параметров
, что означает равенство параметров![]() , т. е. конкурирующие виды в модели Вольтерра не только занимают одну и ту же экологическую нишу, но являются идентичными.
, т. е. конкурирующие виды в модели Вольтерра не только занимают одну и ту же экологическую нишу, но являются идентичными.
В. Вольтерра (Volterra, 1931) построил интересную модель конкуренции внутри одной популяции, разделив ее на две группы, половозрелую численностью x1 и молодь численностью x2. Если в предыдущих моделях предполагалось, что конкуренция уменьшает коэффициенты размножения (роста численности) популяций, то здесь предполагается, что из-за наличия конкуренции увеличивается смертность особей. Уравнения модели имеют вид:
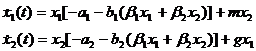 , (1-29)
, (1-29)
где m и g – соответственно коэффициент перехода особей из группы молоди в группу взрослых и коэффициент рождаемости. Параметры a и b –постоянные величины, которые характеризуют смертность от разных причин – не связанную с конкуренцией и связанную с ней соответственно. В. Вольтерра показал, что такая система имеет одну точку равновесия, и решение стремится к ней асимптотически.
Обобщением модели конкуренции Вольтерра является предложенная А. Рисайо и Дж. Ричардсоном (Rescigno and Richardson, 1965) модель конкуренции n видов-потребителей за m типов пищи, которые в разной степени поделены между потребителями. Система уравнений имеет вид:
![]() . (1-30)
. (1-30)
Здесь γij – коэффициент использования j-го корма i-ым видом, Fj – количество пищи j, съедаемое всеми видами в единицу времени. Эта функция положительная и неограниченно возрастает вместе с аргументами. Авторы показали, что (n-m) видов обязательно погибнут (если n>m), и самое большое число видов, которые смогут сосуществовать в этой среде, равно m. К тому же выводу приходит и Хаусман (Haussmann, 1973).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
