
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Исходными материалами (для гл.1) являются разработанные В. Вольтерра (Volterra, 1931) модели взаимодействующих популяций по типу «хищник-жертва» и «конкуренция», а также модели конкуренции видов, предложенные Г. Гаузе (1933,1935). На основе этих моделей нами построена новая модель трофической цепи, в которой потоки вещества с каждого трофического уровня поступают в косную компоненту, а особи первого трофического уровня питаются за счет вещества косной компоненты, а также построены и некоторые другие простые модели сообщества.
Основой для построения многовидовой динамической продукционной модели (гл.3 и 4) стала продукционная модель одной промысловой популяции в состоянии равновесия М. Шефера (Schaefer, 1954). При определении параметров нашей многовидовой модели использованы данные промысловой статистики с 1974 г. по 1988 г. международного промысла в районе Намибии (Статистические сведения…, 1975-1984; ICSEAF, 1976-1985; ICSEAF, 1983), а именно:
· временные ряды общих международных уловов южноафриканской ставриды (Trachurus tr. capensis) и капской мерлузы (Merluccius capensis) района Юго-Восточной Атлантики (Bulgakova and Kizner, 1986; Булгакова и Кизнер, 1985; 1987а), взятых всеми странами и всеми промысловыми комплексами,
· ряды уловов и промыслового усилия на промысле ставриды, в качестве первого стандартного промыслового комплекса выбрано сочетание советского судна типа БМРТ и разноглубинного трала,
· аналогичные данные выбранного в качестве второго стандартного промыслового комплекса на промысле мерлузы в том же районе – советского судна типа БМРТ, оснащенного донным тралом.
Методы стандартизации промыслового усилия и предварительной обработки рядов данных приведены в гл.3 и 4. Оценка параметров модели проводится методом наименьших квадратов.
Исходной моделью для многовидового анализа сообщества Баренцева моря (гл.5 и 6) была когортная многовидовая модель MSVPA (Sparre, 1984; Gislason and Sparre, 1987; Sparre, 1991) в виде программного пакета в среде «Фортран», которая затем была так модифицирована нами и Х. Гисласоном (Дания), чтобы появилась дополнительная возможность использовать переменные параметры (переменные по годам рационы хищника и «остаточная смертность», переменная по сезонам). Эта модель требует больших массивов входной информации. Уловы каждого из четырех видов моделируемого сообщества и их возрастной состав для каждого квартала каждого года из интервала 1980-2006 гг. рассчитывались на основе суммарных международных уловов, долей национальных уловов в общем вылове и возрастной структуры национальных уловов. Но часто возникают сложности с получением информации по возрастному распределению квартальных уловов, поскольку страны-участницы представляют в ИКЕС возрастной состав уловов в целом за год. При расчете возрастного состава квартальных уловов трески до 1994 г. величина международного вылова распределялась по возрастам согласно процентному составу российских квартальных уловов, а начиная с 1994 г. – по данным российской и норвежской промысловой статистики. Распределение международных уловов сельди по кварталам получено из ПИНРО, возрастной состав уловов (в долях) взят из отчетов Рабочей группы ИКЕС (ICES 1996b; 1997b), т. е. его квартальная изменчивость не принималась в расчет. Данные по уловам мойвы и их возрастному составу также получены из ПИНРО. Для креветки Баренцева моря возрастной состав уловов рассчитан по размерному составу уловов с помощью условно принятого в ПИНРО постоянного соответствия размерных и возрастных групп (табл. 6.2).
Входной информацией для MSVPA являются также данные по средней массе одной особи по возрастам, годам и кварталам, как в уловах, так и в запасе – это так называемый «вес особей в море». Данные по массе одной особи в запасе, переменные по кварталам, важны при оценке количества потребленной пищи. Источником такой информации являются размерно-возрастные ключи, полученные из ПИНРО. Для младших и для самых старших возрастных групп они часто не представительны. Удалось найти переменные по годам и кварталам данные по массе особи только для трески и мойвы. Для сельди масса особи по возрастным группам взята одинаковой для всех кварталов года, причем масса особи в уловах и в запасе считается одинаковой. Для креветки приняты неизменные значения массы особи по годам и кварталам (последняя строка табл. 6.2). Отметим, что в данной модели не учитывается тот факт, что из-за избирательности питания хищников средняя масса особи одной и той же возрастной группы жертв в желудках хищников может отличаться от средней их массы в запасе.
Для каждого вида задаются среднемноголетние значения доли половозрелых особей по возрастным группам (огивы половозрелости) (табл. 6.3). Данные для трески, мойвы и сельди рассчитаны по материалам Рабочих групп ИКЕС (ICES 1994; 1996a; 1997b;1998). Для креветки половозрелая часть запаса не имеет большого значения и носит чисто формальный характер, в силу особенностей биологического развития этого вида.
Для оценки параметров питания трески использованы материалы российско-норвежской компьютерной базы данных (БД) по питанию рыб Баренцева моря, начиная с 1984 г. Нами разработаны специальные алгоритмы и программы, позволяющие при работе с этой БД оценить возраст жертв разных видов в желудках хищников, средний за квартал каждого года вес пищи в желудке и средний состав пищи для каждого возраста хищников. Рационы оценивали по методу Б. Богстада и С. Меля (1992), в котором учитывается разная скорость переваривания разных видов жертв в желудке трески, причем скорость переваривания оценена экспериментально именно для трески Баренцева моря (dos Santos, 1990).
Для улучшения качества массивов данных по питанию (чтобы учесть неравномерное распределение трески по ареалу), а также для экстраполяции и интерполяции данных и заполнения пробелов в данных по средней массе пищевого комка и по составу пищи применена методология кригинга (пакет программ WINSURF). Для определения параметров кригинга строились вариограммы с помощью программы GEO-EAS.
Настройка модели MSVPA (гл.6) для трески и сельди проводили на результаты одновидовых оценок запасов (ICES, 1997; 1997b; 1998), при этом искали максимум коэффициента корреляции между рядами численности возрастной группы популяции, полученными по одновидовой и по многовидовой моделям. Настройку мойвы и креветки проводили на результаты учетных съемок.
За основу модели формирования пополнения трески (гл.7) взята модель запас-пополнение У. Рикера (Ricker, 1954), учитывающая смертность молоди от каннибализма. Для усовершенствованной нами модели пополнения трески Баренцева моря популяционная плодовитость трески рассчитана по методу М. Бондаренко и др.(2003), а данные по среднемесячной температуре воды в слое воды 0-200м на станциях 3-7 на Кольском разрезе с разрешения специалистов ПИНРО взяты с сайта ПИНРО. Температурные аномалии рассчитаны по отношению к среднемесячным температурам, оцененным для периода с 1951 г. по 2000 г.
Для построения сценарных моделей трески и сельди исходными данными являются результаты прогонов одновидовых моделей на соответствующих рабочих группах ИКЕС (ICES, 2003; 2005; 2007; 2008).
Глава 1. Математические модели динамики многовидового сообщества и исследование их устойчивости
1.1. Общая блок-схема экосистемы и понятие устойчивости экосистемы
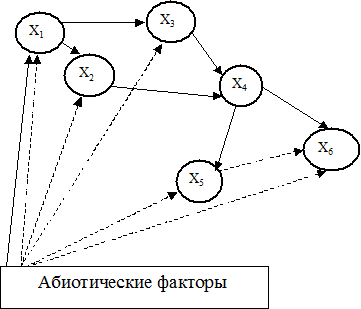
Рисунок 1.1. Общая блок-схема модели экосистемы. Сплошные линии означают потоки вещества или энергии, пунктирные – функциональные связи. Кружки – популяции разных видов.
Представим структуру экосистемы (ЭС) в виде графа, где узлы соответствуют популяциям, а ребра – энергетическим связям между ними (рис. 1.1). В данной схеме сплошные линии означают потоки вещества или энергии, а пунктирные – функциональные связи (некоторые авторы называют их информационными (Меншуткин, 1971).
Возможны два подхода к изучению такой ЭС. Можно, не углубляясь в строение каждой популяции, изучать влияние межпопуляционных связей на динамику ЭС. Условно оборвав эти связи, можно подробно изучать структуру отдельной популяции: при разной степени детализации учитывать половую и возрастную структуру популяции или даже качественный состав биомассы одной особи (содержание в ней белка и жира) (Булгакова и др., 1973), влияние кормовой базы на популяцию (Булгакова, 1973, 1978), процессы естественной и промысловой смертности и размножения, влияние абиотических факторов на динамику различных процессов и т. д. Примерная блок-схема возможной модели популяции приведена на рис. 1.2.
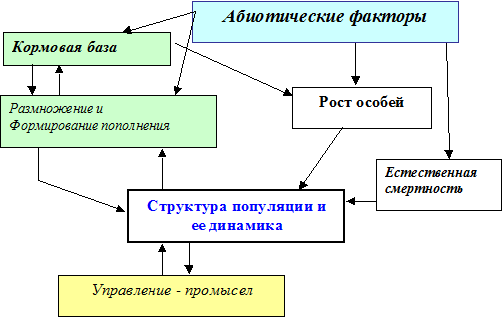
Рисунок 1.2. Блок схемы одной популяции
Подробное описание моделью всех этих факторов может привести к настолько сложной модели, что ее невозможно будет исследовать аналитически. Тем более, при одновременном учете и межпопуляционных связей и структуры каждой популяции приходится строить портретную модель типа модели пелагических рыб озера Дальнего, разработанной , и (1969). Упомянутая модель представляет собой тот редкий случай, который привел к успеху. Такую подробную модель сложно обеспечить необходимой информацией, а поскольку каждый параметр модели определяется с некоторой ошибкой, то при большом количестве параметров ошибки накапливаются, и потому возникает так называемое «проклятие неопределенности», когда надежность выводов, получаемых на основании такой модели, очень низка.
Другой подход, позволяющий учитывать и межвидовые связи и возрастную структуру популяций, реализован в моделях класса MSVPA, которые будут рассмотрены в последующих главах настоящей работы.
В данной главе рассматриваем целый ряд простых моделей сложных экосистем, а именно таких моделей, которые допускают аналитическое исследование. Популяции считаются однородными, при этом изучается влияние межпопуляционных связей разного типа на динамику популяций. Эти модели представляют собой системы обыкновенных дифференциальных уравнений и описывают изменение во времени численности или биомассы каждой компоненты сообщества. Даже для таких простых моделей чаще всего невозможно найти решение в общем виде, но если задать значения модельных параметров, можно численно проинтегрировать системы уравнений и получить решение, описывающее динамику системы.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
