
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Вариант 2). Если a1/2>D/a2 (т. е. M<a1a2/b<2M), то точка S, соответствующая уровню MSY для “изолированной” популяции, лежит за границей допустимой области управления. Это означает, что попытки регулировать первую популяцию с помощью критерия MSY для изолированной популяции приведут к исчезновению хищника (т. е. к разрушению сообщества) даже при отсутствии изъятия хищников! Этот случай – самый опасный для многовидовой системы запас-промысел.
При этом варианте соотношения между параметрами наибольшее значение устойчивого вылова первого вида достигается в граничной точке М (F1= D/a2 и F2=0), лежащей ниже точки S и равно DM/a22. Чтобы не разрушить ЭС, следует выбрать существенно меньшее значение F1 .
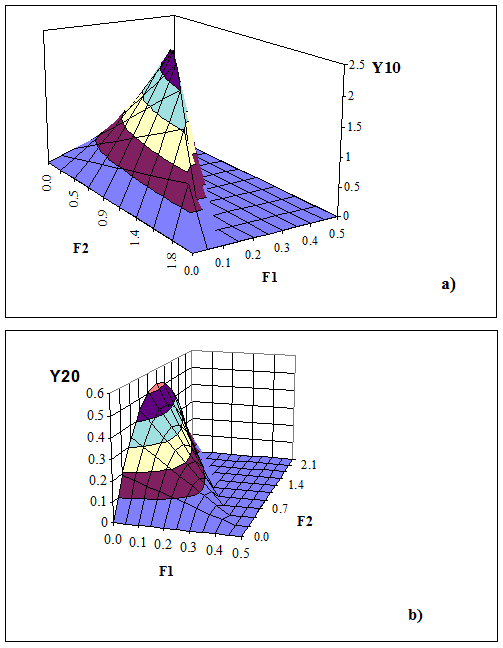
Рисунок 2.2. Равновесный улов вида-жертвы Y10 (a) и вида-хищника Y20(b) в области допустимого управления промыслом для варианта 1 соотношения параметров модели (2-1). На верхнем и нижнем рисунках разное расположение осей координат.
2.1.2. Стратегии специализированного промысла популяции хищников
В устойчивом состоянии сообщества вылов второго вида за год равен
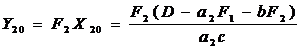 (2-7)
(2-7)
При фиксированном F1 максимум вылова второго вида достигается при
F2 = (D-a2F1)/2b , (2-8)
т. е. на прямой ML, которая является медианой угла OMN и лежит внутри допустимой области управления. Подставив (2-8) в (2-7), получим зависимость Y20 от промысловой смертности вида жертв F1:
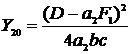 .
.
Это уравнение параболы с вершиной в точке M (рис. 2.1) (F1=D/a2,F2=0), которой соответствует нулевой равновесный улов хищников (рис. 2.2b).
Таким образом, при любом фиксированном значении F1 линия ML определяет уровень промысловой смертности хищников F2MSY, и максимальный вылов хищников зависит от промысловой смертности вида жертв. При этом получаем ожидаемый вывод: максимальное значение вылова популяции хищников можно получить на границе допустимой области – в точке L (F1=0 , F2 = D/2b), т. е. при отсутствии промысла популяции жертв. Точка L может быть выбрана в качестве целевого биологического ориентира при специализированном промысле популяции хищников. При предосторожном подходе следует выбрать точку, лежащую на отрезке ON левее L, например, точку Lpr, в которой F2 равно 0.9D/2b.
На рис. 2.2 построены функции Y10 и Y20 на плоскости аргументов {F1,F2}, параметры модели (2-1) выбраны произвольно, но так, чтобы выполнялось условие варианта 1:
a1 = 0.5; a2 = 5*10-11; b = 10-11; c = 5*10-10; M = 0.2; D = 2.3*10-11..
Дж. Колли и П. Спенсер (Collie and Spencer, 1994) применяли в модели хищник-жертва более сложную функцию для описания процесса хищничества. Если в системе уравнений (2-1) скорость потребления пищи хищником пропорциональна численности жертв a2x1, то эти авторы применяют функцию gx12/(A2+x12), учитывающую существование насыщения (максимального рациона) хищника, которое начинается с уровня численности жертв A. Кроме того, эти авторы представляют естественную смертность хищника в виде квадратичной функции Dx12, учитывая таким образом внутривидовую конкуренцию хищника и (или) каннибализм. Уравнения модели имеют вид
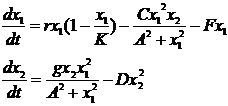 (2-9)
(2-9)
Полученная система уравнений в зависимости от значений параметров модели имеет одну или две устойчивых точки равновесия в положительной области (не считая нулевой), а между ними иногда существует точка неустойчивого равновесия. Очень интересны результаты исследования этой модели: при изменении параметров, зависящих от факторов среды (например, если смертность хищника зависит от температуры), популяции могут “скользить” от одного состояния равновесия к другому. Переход от верхнего уровня равновесия к нижнему может происходить резко также и при переходе коэффициента промысловой смертности жертвы через некоторый порог.
Следует иметь в виду, что в данной главе рассмотрен простейший вариант системы хищник-жертва, когда хищнику доступен единственный вид жертв (пищи). В природных экосистемах обычно существует несколько популяций, которые могут служить пищей для одного вида хищников, при этом снижается опасность исчезновения хищников как популяции из-за ведения нерационального промысла вида жертв. Известно, что при коллапсе мойвы Баренцева моря в середине 80-х годов треска мигрировала в восточные районы моря, переключаясь на питание других видов жертв, в основном, на сайку. При этом ухудшились рост и жизнестойкость трески из-за недостатка запасов липидов в ее печени, но популяция сохранилась.
Включение в модель альтернативной жертвы (Spencer and Collie, 1995) делает модель более реалистичной, но значительно усложняет анализ. Во второе уравнение системы (2-9) в слагаемое, описывающее рост популяции хищников, эти авторы включили еще одно слагаемое, учитывающее биомассу альтернативных жертв. Теперь при стремлении x1 к нулю численность хищников стремится к положительной величине. На рис. 2.3 приведены примеры расположения изоклин для видов хищников и жертв этой модели. Видно, что с ростом F1 понижаются значения численности жертв в состоянии равновесия x10. При повышении F2 область параметров, для которой существует нижнее равновесное состояние жертвы, сужается, т. е. промысел хищников играет роль стабилизатора флюктуаций популяции жертв, способствуя приближению ее численности к верхнему уровню равновесия.
Область допустимого управления для системы (2-1) (рис. 2.2) ограничена прямыми линиями. В более сложных моделях эти линии могут иметь более сложную форму.
Рассмотрим теперь модель хищник-жертва (2-10), предложенную Р. Мэем с соавторами (May et al., 1980) и примененную (1996) к совместному промыслу усатых китов и криля в Антарктике:
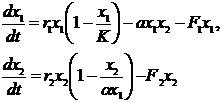 (2-10)
(2-10)
Здесь индексы 1 и 2 относятся к запасам криля и усатых китов соответственно. В данной модели процесс взаимодействия хищника и жертвы описан не так, как в нашей модели (2-1): коэффициенты естественного прироста численности обеих популяций r1, r2 приняты постоянными величинами; а ёмкость среды хищников (китов) считается пропорциональной величине запаса криля, то есть K2=αx1. Параметр a –коэффициент, характеризующий влияние запаса криля на запас китов.
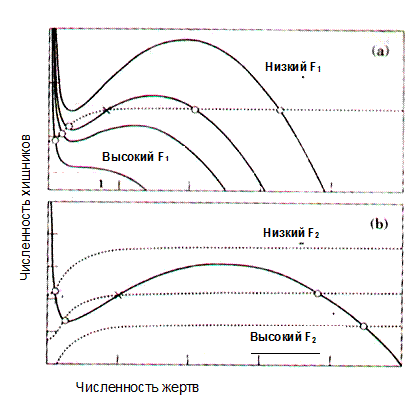
Рисунок 2.3. Фазовая плоскость {x1,x2} и семейство изоклин для вида жертв (пунктирные линии) и вида хищников (сплошные линии) (по Spencer and Collie, 1995): влияние промысловой смертности жертв F1 на положение точек равновесия (a); влияние промысловой смертности хищника F2 на положение точек равновесия системы (b).
Для промысловой системы (2.10) координаты точки равновесия равны
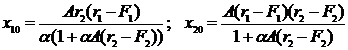 . (2-11)
. (2-11)
Здесь A=αK/r1r2. Из формулы (2-11) видно, что область допустимого управления для этой системы ограничена условиями F1< r1, F2< r2, т. е. представляет собой прямоугольник, в отличие от области, приведенной на рис. 2.1. Анализ показывает (Бородин, 1996; Булгакова и др., 2002; Многовидовой анализ…2001), что при фиксированном F2 улов криля будет максимальным при F1=0,5r1. С ростом интенсивности промысла китов (F2) MSY криля растет при снижении запасов и величины добычи китов. Если параметр F2 достигнет уровня r2, то киты исчезнут, а MSY для криля будет наибольшим.
Рассмотрим теперь равновесный вылов китов Y10 =F2x20. Этот вылов равен нулю при F2=0 и при F2=r2 и достигает максимума внутри этого интервала, точнее – внутри интервала (r2/3, r2/3+1/3a). Положение максимума (соответствующее ему значение F2) не зависит от F1, но величина максимального уравновешенного улова (MSY) китов линейно возрастает с уменьшением F1 до нуля. Эти выводы совпадают с выводами, полученными для нашей модели хищник-жертва (2-1).
2.1.3. Регулирование двухвидового промысла системы хищник-жертва с учетом экономического критерия
Проблема оптимального управления сообществом в целом приводит к необходимости рассмотреть вопрос максимизации экономической прибыли двухвидового промысла в равновесном режиме за единицу времени (например, за год), варьируя значения F1 и F2 внутри допустимой области управления OMN. В простейшем случае, если независимо ведутся два специализированных промысла, экономическая прибыль выражается в виде функции
P(F1,F2) = A1F1(M + F2)/a2 + A2F2(D-a2F1-bF2)/ca2 -B1F1-B2F2 (2-11)
Параметры A1 и A2 - стоимость единицы вылова I и II популяции соответственно, B'1 и B'2 - затраты на единицу промыслового усилия E1 и E2 соответственно. Тогда при предположении пропорциональности коэффициента промысловой смертности промысловому усилию Fi=qiEi (i=1,2) и при постоянных коэффициентах qi можно считать, что B1=B'1/q1 и B2=B'2/q2. Отметим, что в функции прибыли (2-11) опущены другие категории затрат на промысел, например, постоянные затраты, не связанные с промысловым усилием, или зависящие от величины вылова.
Уравнение (2-11) перепишем в виде полинома
P(F1,F2) = k1F1 + k2F2 - k3F1F2 -k4 F22 (2-12)
k1 = A1X10 - B1 ;
k2 = A2X20 - B2;
k3 = - A1/a2 + A2/c ;
k4 = A2b/ca2.
Здесь X10 и X20 - координаты точки равновесия неэксплуатируемой (неуправляемой) системы – (согласно уравнению 2-2), следовательно, k1 и k2 – прибыль, получаемая на единицу коэффициента промысловой смертности в этой точке равновесия. Если промысел в этой точке рентабелен, k1 и k2 - положительные константы. Знак коэффициента k3 может быть любым, например, если A2/c>A1/a2, то популяция хищников является более ценной, чем популяция жертв, тогда k3>0, и наоборот.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
