
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Конечно, всегда следует помнить, что полученное решение описывает динамику модельных представлений об изучаемой ЭС, а не самой ЭС. Сопоставление решения уравнений модели с динамикой реальной системы является в некоторой степени проверкой адекватности модели. Но получить истинную динамику природной экосистемы, с которой можно было бы сравнить рассчитанную по модели динамику, практически невозможно. Так, проводимые часто учетные съемки, траловые или акустические, или какие-нибудь иные определяют численность запасов с очень высокой ошибкой, и в лучшем случае дают только индексы численности или биомассы.
Одним из способов анализа адекватности аналитической модели является исследование на устойчивость ее решения. Как правило, реальная ЭС существует достаточно долго (в масштабах времени наблюдений). Если решение показывает, что одна из компонент модели через некоторое время исчезает (ее численность стремится к нулю), т. е. нарушается структура экосистемы, то следует считать модель не соответствующей оригиналу. Аналогично, если численность какого-нибудь вида сообщества со временем неограниченно возрастает, это тоже говорит о несоответствии модели моделируемой ЭС, поскольку в природе всегда существуют факторы, ограничивающие (лимитирующие) рост численности. Согласно концепциям термодинамики «любая замкнутая система с протекающим сквозь нее потоком энергии (будь то биосфера или маленькое озеро) с большой вероятностью развивается в сторону некоторого устойчивого состояния, и в ней с необходимостью должны вырабатываться саморегулирующие механизмы» (Свирежев и Логофет, 1978).
Можно рассматривать устойчивость экосистем разной степени иерархии, разного пространственного и временного масштаба (Свирежев и Логофет, 1978; Свирежев, 1987).
1. Устойчивость понимается как неизменность во времени целого географического региона1.[1]Экосистема региона или ландшафта может включать в себя несколько различных ЭС, слабо связанных друг с другом.
2. Устойчивость рассматривается как сохранение числа видов в сообществе в течение длительного времени. Эту устойчивость можно назвать структурной устойчивостью.
3. Сообщество считается устойчивым (стабильным), если численности составляющих его популяций не испытывают резких колебаний.
Вопросы устойчивости региональных ЭС в данной работе не рассматриваются, а второе и третье определение вполне соответствуют понятиям математической устойчивости системы дифференциальных уравнений. Теория математической устойчивости решений таких систем уравнений разработана, но надо помнить, что она относится к математическим моделям, а не к реальным природным экосистемам.
Устойчивость по (1950) в применении в ЭС трактуется следующим образом. Пусть моделируемое сообщество имеет стационарное (равновесное) решение, которое не является тривиальным. Равновесному состоянию соответствует точка на фазовой плоскости. Нетривиальность решения означает, что для этого стационарного состояния численности всех видов сообщества не равны нулю. Если сообщество находится строго в стационарной точке, то численности всех входящих в него популяций не меняются во времени, поскольку их производные по времени в модели равны нулю. Устойчивость по Ляпунову этого решения означает, что при малых отклонениях системы от состояния равновесия система стремится к нему вернуться. Таким образом, существование устойчивого равновесия обеспечивает и структурную устойчивость ЭС (сохранение всех видов сообщества).
В то же время сохранение всех видов сообщества возможно и в случае отсутствия равновесного состояния системы, если траектории изменения ее состояния лежат в положительной области и ограничены сверху и снизу. При этих условиях ни один вид не вымирает, т. е. сохраняется структура сообщества, но условие устойчивости по Ляпунову не выполняется. Такая устойчивость ЭС соответствует понятию устойчивости по Лагранжу.
По-видимому, в природных ЭС в процессе эволюции происходят переходы от одного устойчивого состояния равновесия к другому через неустойчивые состояния и бифуркации (Свирежев, 1987). При исследовании же динамики ЭС в данной работе имеются в виду масштабы времени, гораздо меньшие, чем время, за которое происходят заметные эволюционные процессы. В настоящей главе используется понятие устойчивости по (1950).
В этой главе рассмотрены модели сообществ с учетом только биологических экосистемных (эндогенных) факторов, но особенно важно исследование устойчивости эксплуатируемой ЭС, при котором определяются такие значения степени эксплуатации (или вылова), при которых не нарушается ее устойчивость. Эти вопросы рассмотрены позже (гл. 2).
1.2. Исследование моделей трофических цепей и их устойчивости
В данном разделе рассматриваются математические модели (ММ) экосистем такого типа, когда особи каждого последующего звена (численностью xi) питаются только особями предыдущего звена (xi-1), а сами являются пищей только для особей последующего (xi+1), здесь i =1,2,…n; n – число звеньев. Такая структура называется трофической цепью или трофической пирамидой (рис. 1.3).
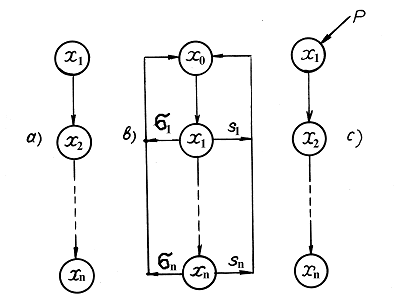
Рисунок 1-3. Блок схемы трех трофических цепей, соответствующих модели Вольтерра (Volterra, 1931) (а), модели n видов с обратной связью на косную компоненту типа «запас» (b) и модели n видов с косной компонентой типа «поток» (c) (по Булгаковой,1966)
Если полагать, что каждое звено цепи соответствует одной популяции, то ЭС с такой структурой редко встречается в природе, особенно при большом значении n. Если же полагать, что каждое звено представляет собой целый трофический уровень, который может включать несколько популяций разных видов, то ЭС или биогеоценозы такой структуры распространены довольно широко. Таким образом, в данном разделе не рассматриваются трофические сети, т. е. структуры, в которых особи некоторых звеньев могут питаться особями более чем одного из предшествующих звеньев.
При построении ММ считаем трофический уровень однородной совокупностью особей и пренебрегаем какими-либо взаимоотношениями между его составляющими видами. Под xi понимаем численность i-го уровня или суммарную его биомассу или количество некоторого вещества (или его энергетического эквивалента), заключенного в особях этого уровня. Например, если рассматривать пелагическое сообщество, I-й трофический уровень соответствует фитопланктону (продуцентам), II-й – зоопланктону-фитофагу, III-й – хищному зоопланктону и т. д.
Рассмотрим влияние абиотических факторов разного типа на устойчивость ценоза.
1.2.1.Модель трофической цепи В. Вольтерра
В. Вольтерра (Volterra, 1931; Volterra and d’Ancona, 1935) при построении ММ трофической цепи не учитывал влияния на абиотических факторов на биологическое сообщество, в основу модели положены следующие гипотезы:
1. Величины xi - положительные непрерывные функции;
2. Изменение величины xi за время Δt обусловлено тремя причинами: рождением новых особей (Δ i1), естественной смертностью (-Δi2) и убылью за счет пожирания хищниками (i+1)-го уровня (-Δi3), то есть
Δxi= (Δ i1-Δ i2- Δi3) Δt.
2a) Величина Δi1 для малого Δt пропорциональна числу встреч между особями i-го и (i-1)-го уровней, т. е. пропорциональна произведению числа особей (или биомассы) i - го уровня и количества пищи для него: Δi1 =εixixi-1, где εi- постоянная положительная величина, имеющая смысл скорости размножения особей i-го уровня; полагаем, что для первого уровня пища в изобилии, потому Δ11=ε1x1.
2б) Величина Δi2 для малого промежутка времени Δt пропорциональна численности i-го уровня и равна sixi, где si -положительная константа, имеющая смысл мгновенного коэффициента естественной смертности.
2в) Величина Δi3 для малого Δt пропорциональна числу встреч между особями i-го и (i +1)-го уровней и равна Δi1 = δi+1xixi+1, где δi+1 – положительная константа, имеющая смысл коэффициента активности хищников (i+1)-го уровня.
Система дифференциальных уравнений модели имеет вид:
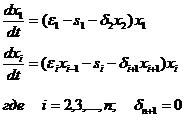 (1-1)
(1-1)
Модель (1-1) является одной из рассмотренных В. Вольтера моделей биологических сообществ, но именно эту модель в дальнейшем называем моделью Вольтерра. Блок-схема такой модели приведена на рис. 1.3a. Координаты стационарной точки системы определяются приравниванием правой части уравнений нулю и решением полученной при этом системы линейных алгебраических уравнений. Понятно, что биологический смысл имеют только такие случаи, когда координаты точки равновесия являются положительными величинами.
Исследование этой системы уравнений для n=2 детально проведено В. Вольтерра (Volterra, 1931; перевод на русский – Вольтерра, 1976), оно довольно громоздко и здесь не приводится. Решение имеет вид незатухающих колебаний возле точки равновесия, причем период малых колебаний относительно этой точки равен T=2π/√(ε1ε2).
Для более общего случая при n>2 там же показано, что точка равновесия системы (1-1) может (при некоторых соотношениях между параметрами) существовать в положительной области (т. е. xio>0 для i=1,2,.., n) только при четном n. При этом при небольших начальных отклонениях от точки равновесия биомасса каждого вида меняется с незатухающими колебаниями возле точки равновесия, т. е. находится на границе устойчивости.
Исследование системы (1-1) на устойчивость стационарной точки проведено нами (Булгакова, 1966) по первой методе (1950). Схематически такое исследование состоит из следующих этапов:
1. Решение системы алгебраических уравнений, полученных после приравнивания нулю правых частей уравнений (1-1), определяет точку равновесия сообщества {xio}. Определяются условия, при которых эта точка лежит в положительной области. Эти условия выражены в виде соотношения между параметрами популяций сообщества.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
