
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
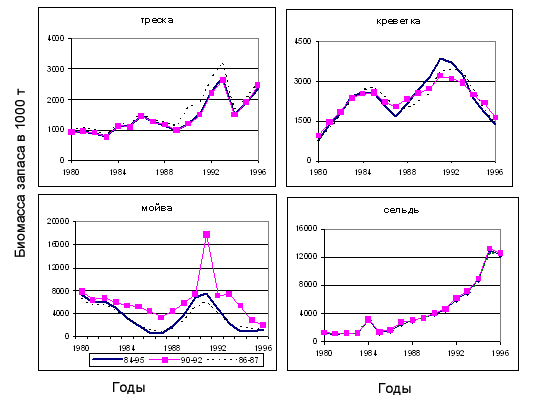
Рисунок 6.24. Оценки биомассы запасов при традиционном варианте настройки MSVPA при разных наборах данных по составу желудков: все имеющиеся данные за период с 1984г. по 1995 г.; годы с высоким уровнем мойвы 1990-1992; годы с низким уровнем мойвы 1986-1987.
Таким образом, показано, насколько значительными могут быть отклонения (ошибки) оценок многих популяционных характеристик, полученных с помощью модели MSVPA, если мы располагаем данными по составу желудков за короткий период. Особенно велики эти отклонения для видов, все возрастные группы которых являются объектом хищничества (как мойва и креветка). Оценки биомассы или численности видов с многовозрастной структурой, из которой лишь младшие возрастные группы подвержены хищничеству (такие как треска и сельдь), меньше зависят как от выбранного набора данных по питанию, как и от разных версий модели.
Развивая многовидовую модель сообщества Баренцева моря, и (1999) включили в сообщество еще одного хищника-пикшу, два дополнительных вида жертв – молодь пикши и сайку и двух "внешних" хищников – гренландского тюленя и кита-малого полосатика. Из-за недостатка информации по млекопитающим было сделано предположение, что их численность и рационы не меняются по годам.
В начале 2000-х годов численность тюлений и китов Северо-Восточной Атлантики, влияющих на многие виды рыб экосистемы, была так велика, что не позволяла наращивать вылов рыб. Недостоверные данные по численности видов млекопитающих и по их питанию (спектру питания и величине рационов) не позволяют получить надежных оценок этого влияния.
Показано (Коржев и Долгов, 1999), что роль пикши как хищника значительно меньше, чем трески. Больше всего пикша съедает мойвы и креветки, в отдельные годы до 10 000 т., но в основе ее питания лежит нерыбная пища. Роль молоди пикши в рационе трески мала – менее 1 %.
Доля сайки в питании трески менялась, достигая 5% (по биомассе) в 1995 г. В среднем в последние годы расчетного периода треска потребляла около 200 000 т сайки в год, при этом запас сайки в эти годы меняется от 400 000 до 1 000 000 т. (Коржев, Долгов, 1999).
На рис. 6.25 в виде кумулятивных функций показана динамика потребление треской разных видов, в тоннах. Поскольку потребленная треской биомасса разных видов жертв различается на порядок величин, рисунок разбит на два, и оси ординат этих рисунков даны в разном масштабе. Так что для каждого года суммарная биомасса пищи, потребленной треской в течение этого года, равна сумме двух верхних функций на этих рисунках. Доля другой пищи в питании трески при таком моделировании составляет от 39 до 69%, т. е. немного уменьшилась при добавлении двух видов жертв.
Оценки влияния млекопитающих следует считать предварительными из-за указанных выше упрощений. Тем не менее, в первом приближении можно сказать, что их влияние на формирование молоди трески велико (рис.6.26) – при возрастании оценок смертности молоди трески от хищничества при включении млекопитающих резко возросли оценки численности трески. Аналогичная картина получена авторами для пикши и сельди. Конечно, в процессе работы с этой моделью следует уточнить оценки остаточной смертности, поскольку параметры М1 были оценены с учетом всех других причин, кроме хищничества трески, в том числе они могли включать и смертность от млекопитающих.
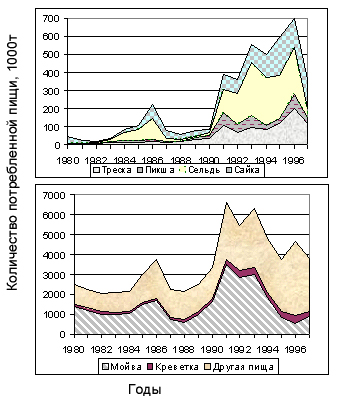
Рисунок 6.25. Потребление шести видов жертв и другой пищи популяцией трески по годам, в т, показанное в виде кумулятивных функций (построено по материалам Коржева и Долгова, 1999).
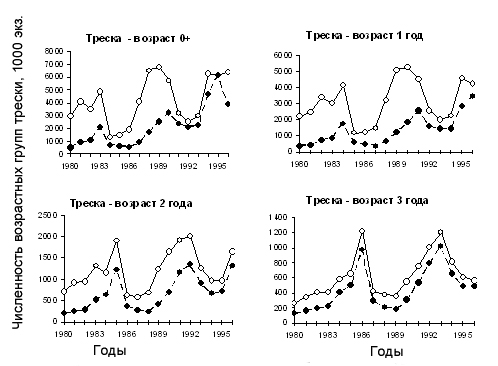
Рисунок 6.26. Динамика численности младших возрастных групп трески, рассчитанная по MSVPA с учетом (пустые кружки) и без учета (заполненные кружки) влияния морских млекопитающих (по Коржеву и Долгову, 1999)
6.5. Анализ чувствительности модели MSVPA
В период работы в Рабочей группе ИКЕС по многовидовым оценкам в Балтике автором был проведен анализ чувствительности трехвидовой (пятикомпонентной) модели MSVPA сообщества Восточной Балтики к терминальным значениям промысловой смертности различных компонент (ICES, 1990). Структура этого сообщества включала один запас хищника-трески, два запаса шпрота (подрайоны 25 и 26+28) и два запаса сельди (подрайоны 25-27 и 28-29). В качестве базового был выбран прогон модели, выполненный этой рабочей группой в 1989 г. В качестве выходных величин выбраны оценки общей биомассы каждого компонента и иногда – количество потребленной биомассы видов жертв.
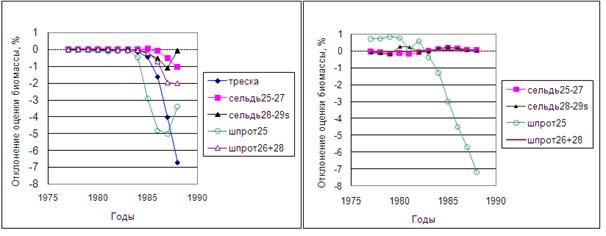
Рисунок 6.27. Анализ чувствительности модели MSVPA на примере сообщества Балтийского моря: относительные отклонения биомассы разных компонентов в % при +10%-ом изменении Fterm хищника-трески (левый график) или Fterm жертвы – шпрота подрайона 25 (правый график).
На левом рис. 6.27 и в табл. 6.10 показано влияние 10%-го увеличения Fterm хищника-трески на оценки биомассы всех видов сообщества. При этом эксперименте оценка биомассы трески в терминальный (1988) год получена заниженной на 6.7%, и при удалении от терминального года в ретроспективу эти изменения уменьшались, уже через 5 лет став пренебрежимо малыми. Занижение оценки биомассы хищника в свою очередь приводит к уменьшению коэффициентов смертности от хищничества видов-жертв, следовательно, и к занижению оценок биомасс всех видов жертв, поскольку расчеты ведутся ретроспективно.
Относительное изменение биомассы каждого запаса жертв меньше, чем хищника, но из запасов жертв наиболее чувствительным оказался шпрот подрайона 25. Этот факт следует учитывать в процессе настройки модели, поскольку именно на оценках этого запаса в первую очередь отразятся неточности определения параметров модели.
Увеличение на 10% коэффициента Fterm одного из видов жертв (в данном случае шпрота подрайона 25) практически не влияет на запас хищника, но занижает оценку запаса самого шпрота на 7%. В результате происходит перераспределение оценок потребленных треской пищевых компонентов, и оценки биомассы других видов жертв несколько завышаются, хотя эти отклонения относительно малы (правый рис. 6.27 и табл. 6.11).
Таблица 6. 10
Влияние отклонений значений коэффициентов Fterm (для 1988 г.) трески на оценки биомассы запасов относительно базового расчета по модели MSVPA сообщества Восточной части Балтийского моря (все в %).
Отклонение Fterm трески, % | Отклонение оценки биомассы запаса в 1988 г.,% | ||||
Треска | Сельдь 25-27 | Сельдь 28-29 | Шпрот 25 | Шпрот 26+28 | |
+10 | -6,7 | -1,1 | -1,1 | -5,0 | -2,0 |
+20 | -12,2 | -1,9 | -2,0 | -8,8 | -3,7 |
+100 | -36,1 | -5,4 | -5,9 | -24,1 | -10,7 |
Отклонение оценки биомассы, потребленной хищниками, % | |||||
+10 | - | -9,1 | -8,3 | -9,6 | -9,3 |
+20 | - | -16,2 | -14,9 | -17,2 | -16,8 |
+100 | - | -47,3 | -45,0 | -48,3 | -49,0 |
+ увеличение показателя; - уменьшение
Увеличение Fterm трески вызывает отрицательные отклонения всех показателей.
Таблица 6.11
Влияние отклонений Fterm (в 1988 г.) шпрота подрайона 25 на оценки биомассы запасов относительно базового расчета по модели MSVPA сообщества Балтийского моря, (в %).
Отклонение Fterm шпрота 25, в % | Отклонение оценки биомассы запаса в 1988 г. | |||
Сельдь 25-27 | Сельдь 28+29 | Шпрот 25 | Шпрот 26+28 | |
+10 | +0,04 | +0,01 | -7,2 | +0,02 |
+100 | +0,16 | +0,06 | -40,7 | +1,0 |
Отклонение оценки биомассы, потребленной хищниками в терминальном 1988 г. | ||||
+10 | +0,04 | +0,01 | -4,2 | +0,02 |
+100 | +0,76 | +0,1 | -44,2 | +0,32 |
Расчеты чувствительности, приведенные выше, относятся к прогону модели в 1989 г., когда использовались данные по составу желудков трески только за один год (1983). По-видимому, расширение базы данных по составу желудков должно повлиять на результаты моделирования и на чувствительность модели.
Модель MSVPA была применена к сообществу девяти видов рыб банки Джорджес для периода с 1978 г. по 1992 г. (Tsou and Collie, 2001). Из девяти видов шесть описаны детально – как "включенные в модель" (атлантическая треска, серебристый хек, атлантическая скумбрия, пикша, атлантическая сельдь и желтохвостая камбала), два вида (колючая акула и зимний скат) рассматривались как внешние хищники и один – как внешняя жертва (песчанка). Это означает, что для трех последних видов оценки численности получены вне модели, а именно из учетных съемок. Анализ показал (Tsou and Collie, 2001), что для этой ЭС наиболее высока чувствительность модели к величинам общего рациона хищников, следующий по важности фактор – терминальные значения промысловой смертности. Если входные параметры модели менялись в интервале ±25%, то выходные – в пределах ±10%. Получено, что смертность от хищников видов жертв положительно связана с изменением их собственных Fterm и отрицательно связана с собственными значениями остаточной смертности M1 вида жертв. Это вполне ожидаемый результат, поскольку высокие значения Fterm и низкие значения M1 приводят к более низким оценкам численности данного вида и в результате – к завышению оценок M2. Кроме того, высокое значение M2 может быть и результатом завышенных оценок численности хищников, полученных из-за завышенных оценок Fterm для тех же видов хищников.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
