
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рисунок 6.18. Сравнение результатов расчетов по MSVPA, полученных двумя способами настройки трески и сельди: оценки биомассы нерестового запаса четырех видов, в 1000 т; сплошные линии соответствуют традиционному методу, пунктир с метками – настройке с помощью QISVPA.
Сравнение результатов двух методов настройки демонстрируется на примере полученных по модели оценок нерестовой биомассы каждого из четырех видов (рис. 6.18). Видно, что MSVPA-QISVPA – процедура настройки дает несколько отличные от традиционного способа настройки оценки запаса в последние годы. Различия в оценках запасов быстро падают при удалении от терминального года и практически исчезают, начиная с 1992г., несмотря на то, что в двух процедурах настройки использовались различные оценки как M1, так и Fterm для старших возрастных групп всех расчетных лет, а не только для последнего года. Для сельди при этом получается более сглаженная картина для 1984г. Оценки Fterm(a) для терминального года также несколько отличаются от полученных с использованием «традиционного» способа их оценки.

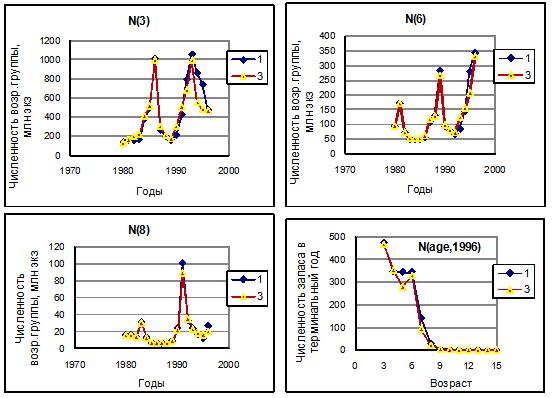
Рисунок 6.19. Сравнение межгодовой динамики оценок численности разных возрастных групп трески, а также распределения по возрастам численности запаса трески в терминальный год, полученных по одновидовому ВПА(1) и после многовидовой настройки MSVPA (3) на результаты одновидового ВПА.
Оценки численности запаса трески, полученные по MSVPA при «традиционной» настройке, практически совпадают с оценками по одновидовому ВПА, как и следовало ожидать (рис. 6.19). После настройки по этому способу коэффициент корреляции между временными рядами численности возрастных групп трески по одновидовым расчетам ВПА и по MSVPA достигал значений 0,95-0,99, для сельди – 0,88-0,99. Коэффициент корреляции между временными рядами биомассы мойвы в начале IV квартала по съемкам и по MSVPA достигал величины 0,90. Столь же высока корреляция оценок биомассы креветки по модели в начале II квартала с результатами весенних съемок. Но поскольку настройка мойвы и креветки велась только по биомассе запаса, различия в возрастной структуре запаса, полученные по MSVPA и по съемкам, остаются существенными (рис. 6.20).
Результаты, полученные при втором методе настройки, получены независимо от одновидовых расчетов и могут считаться более близкими к реальности, поскольку дополнительно учитывают взаимоотношения между видами. Но второй метод настройки требует наличия дополнительной одновидовой модели оценки запаса и трудоемких одновидовых расчетов для каждого из настраиваемых видов и многократных расчетов по MSVPA. Далее в качестве "основного" прогона модели рассматриваем прогон с традиционным методом настройки.
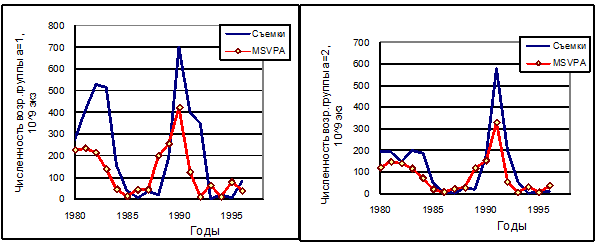
Рисунок 6.20. Сравнение динамики численности двух возрастных групп мойвы (один год – левый рисунок и два года – правый рисунок), оцененных по осенним съемкам на 1 октября и по модели MSVPA (основной прогон).
Сопоставление полученных по модели оценок динамики биомассы запаса, потребляемой хищниками за год биомассы и годового вылова (рис.6.21) показало, что потребленная треской биомасса ее молоди невелика по сравнению с выловом, но на самом деле пресс каннибализма весьма значителен – параметр M2 для молоди трески достаточно высок, особенно для возрастной группы 1 (рис. 6.22). Этот пресс усиливается в годы низкого запаса мойвы и начиная с 1992 г. – при увеличении общего запаса трески.
В основном прогоне модели ярко видна главная роль мойвы в питании трески. При низком запасе мойвы резко усиливается потребление креветки (рис.6.21), трески и сельди (последней – особенно в последние годы при увеличении запаса сельди), а также треска бывает вынуждена в большей степени перейти на ”другую” пищу.
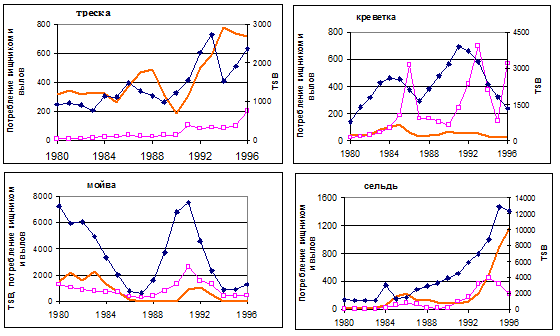
Рисунок 6.21. Результаты основного модельного расчета: для каждого из видов оценки общего запаса (TSB) на начало года – линии с темными ромбиками (для всех видов, кроме мойвы, этим оценкам соответствует правая ось ординат); биомасса, потребленная за год треской – линии с метками в виде пустых квадратиков; годовой вылов – сплошная линия; все в 1000 т.
В целом, результаты применения MSVPA показали, что в течение трех последних исследованных лет (1994-1996гг.) при низкой численности мойвы и высокой численности сельди треска питается главным образом «другой пищей» (до 70 % общей потребленной треской биомассы). В эти же годы креветка, сельдь и мойва дают вклад около 8-10% каждый (рис. 6.23a).
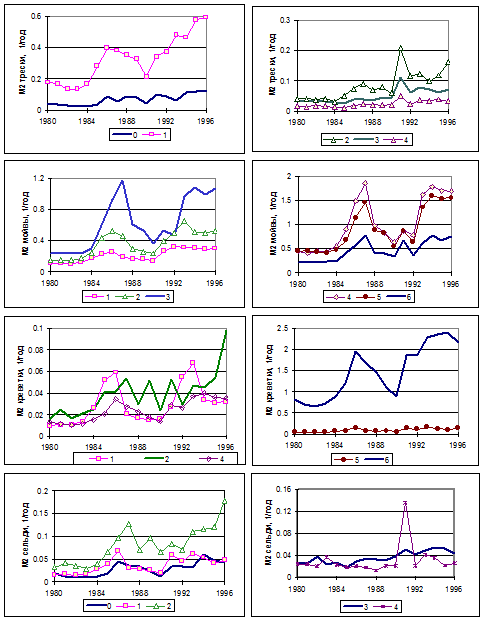
Рисунок 6.22. Результаты основного модельного расчета. Динамика коэффициента смертности от хищничества M2(a) для каждого из четырех видов по годам, отдельно для разных возрастных групп.
Потребление сельди треской в большинстве случаев соизмеримо с ее выловом, причем наибольшие значения M2 получены для двухлетней сельди (рис. 6.22). Потребление мойвы треской во многие годы существенно превышает вылов мойвы. Смертность мойвы от хищника-трески особенно велика для возрастных групп 3, 4 и 5 лет (рис. 6.22) и усиливается для всех возрастных групп в годы с низким запасом мойвы и при росте запаса трески.
Потребление треской креветки после 1984 г. иногда в несколько раз выше объема ее вылова (рис. 6.21). Наибольшие значения коэффициента M2 креветки получены для возраста 6 лет (рис. 6.22), возможно, это результат очень низкой численности этой возрастной группы.
Основная информация, необходимая для моделирования взаимоотношений видов в модели MSVPA (да и во многих других моделях, описывающих многовидовое сообщество), заключена в данных по составу желудков хищников, то есть в данной версии модели – трески. Поскольку получение этой информации является наиболее трудоемкой частью работы по многовидовому моделированию, при отсутствии длинных рядов данных часто приходится ограничиваться данными по питанию за 2-3 года. В течение рассматриваемого периода с 1984 г. по 1995 г. резко менялись численность как трески, так и разных видов ее жертв, особенно мойвы, которая является основным определяющим питание и состояние трески видом.
Чтобы проверить чувствительность модели к выбираемому исследователем набору данных по питанию, проводились прогоны с разными наборами данных по составу желудков, а именно: для всего указанного периода (1984-1995 гг.) (основной прогон), для периода 1986-1987гг., в который мойва была в депрессии, и для периода 1990-1992гг., когда запас мойвы восстановился (рис.6.23). В основном прогоне рационы хищника-трески варьировали не только по сезонам (кварталам), но и по годам, а в остальных прогонах использованы средние за выбранный период рационы, но переменные по кварталам. Во всех прогонах рацион зависел от возраста трески. Для сравнения приводится и вариант оценки запасов со среднемноголетними за весь интервал лет 1984-1995 значениями квартальных рационов (квартальные различия при этом все же учитывались).
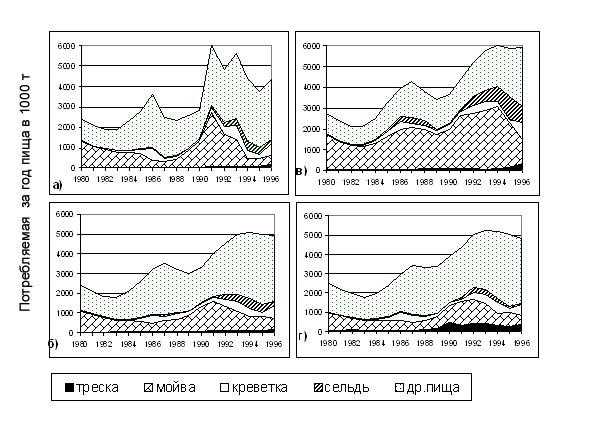
Рисунок 6.23. Кумулятивные графики, демонстрирующие состав и количество потребляемой за год пищи всей популяцией трески (в 1000 т), полученные в результате моделирования для трех наборов данных по составу пищи трески:
(a) взяты все имеющиеся данные за период с 1984г. по 1995 г., рацион трески меняется по годам и кварталам R(a,Y,Q);
(б) данные по составу пищи те же, что в варианте (а), а для каждого квартала рацион рассчитывается как средний за весь период с 1984 по1995 гг. – R(a,Q);
(в) выбраны данные по составу пищи за годы с высоким уровнем мойвы 1990-1992гг., и рацион усреднен для этих лет R(a,Q);
(г) данные взяты в годы с низким уровнем мойвы 1986-1987гг., и рацион усреднен за эти годы.
Рис.6.23а демонстрирует значительную изменчивость потребления треской разных видов жертв и главную роль мойвы в питании трески. Здесь приводится суммарная потребленная биомасса за год, в первом квартале доля мойвы в питании трески еще выше. Видно, что в более поздние годы увеличивается роль сельди и собственной молоди трески в питании трески. Считаем, что в этом варианте расчетов получены наилучшие оценки, поскольку в этом случае использован наиболее полный массив входной информации.
Следующий вариант расчетов (рис. 6.23б) показывает, что в результате усреднения квартальных рационов по годам получается сильно сглаженная динамика потребленной хищником биомассы каждого вида жертв, но основные тенденции при этом сохраняются. Например, в годы с низким запасом мойвы отклонения потребленной биомассы мойвы от основного расчета достигают 80-90%. Для креветки наблюдается та же картина. При этом оказываются сглаженными и колебания численности запасов отдельных видов. Эти машинные эксперименты показывают, что для получения более реальной динамики запасов необходимо включить в модель межгодовую изменчивость квартальных рационов хищника, во всяком случае, это заключение справедливо для бореальной экосистемы.
Если же взять за основу данные по составу желудков за те годы, в которые было много мойвы (рис. 6.23в), то возрастают как оценки суммарной биомассы потребленной пищи (верхняя кривая на этом рисунке), так и доля мойвы в ней. Поэтому и оценки запаса мойвы получаются сильно завышенными (рис. 6.24) по сравнению с основным расчетом, например, для 1991 г. – в 5 раз.
Если взять за основу данные по составу желудков за годы, когда было мало мойвы (рис. 6.23г), то получаем еще более сложную картину. Период с 1986 г. по1987 г. характеризуется не только низким запасом мойвы, но и выделяется составом пищи трески, потому распространение данных по составу желудков этих лет на весь период исследований дает сильные и неожиданные смещения оценок. Усиливается потребление трески и сельди (последней – особенно в последние годы при увеличении запаса сельди), биомасса потребленной молоди трески становится заметной (рис. 6.23г) в течение всего расчетного периода. При таких расчетах получается, что треска вынуждена в большей степени (по сравнению с основным прогоном модели) перейти на «другую» пищу. Оценки биомассы трески в этом варианте расчетов завышены для всех лет, отклонения достигают 40% в 1990 г. и 1993 г. (рис. 6.24). Оценки запаса мойвы занижены, параметр M2 этого вида резко флюктуирует, давая пики в годы низкого уровня запаса. Отклонения запаса креветки – в пределах 20%. Оценки запаса сельди практически не изменились, хотя потребление сельди в одни годы завышено (в 1984 г. - в 1,5 раза), а в другие – занижено.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
