
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
3.2. Многовидовая продукционная модель – равновесный вариант
Поскольку промысловые виды, как правило, существуют не изолированно друг от друга и часто взаимодействуют, эксплуатация одного из видов сообщества может влиять на результаты промысла других, то есть требуется разработка и исследование моделей многовидового промысла, приближенных к реальности. В связи с этим нужно иметь в виду, что чем сложнее модель, чем больше она включает структурных элементов и функциональных связей, тем больше исходных данных требуется для ее реализации и тем детальнее необходимо разбираться, например, в вопросах поведения, питания, возрастной структуры моделируемых видов рыб. В то же время простые модели, верно отражая основные тенденции изучаемых процессов, требуют меньше исходной информации и обычно менее чувствительны к ее качеству.
Подобные соображения, вероятно, сыграли не последнюю роль в том, что развитие прикладных исследований по моделированию многовидовых эксплуатируемых сообществ морских организмов пошло по пути прямого обобщения традиционных одновидовых моделей.
Рассмотрим построенную на базе модели Шефера модель одного промысла, эксплуатирующего одновременно запасы двух биологически не взаимодействующих видов (Horwood, I976):
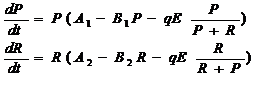 , (3-5)
, (3-5)
где P и R – обилие каждого из видов (численность или биомасса запасов). Это простейшая модель «технологического» взаимодействия двух видов, при котором коэффициенты промысловой смертности каждого вида считаются пропорциональными относительным долям запасов этих видов в суммарном запасе. Точнее, величина промыслового усилия распределяется между видами пропорционально их доле в общем запасе при условии, что коэффициенты улавливаемости видов q равны.
Модель двух взаимодействующих однородных популяций, предложенная Дж. Поупом и О. Харрисом (Pope and Harris, 1975), представляет собой систему двух дифференциальных уравнений (3-6), причем популяции могут взаимодействовать по типу хищник-жертва или конкурировать за пищу, а тип взаимодействия зависит от знаков параметров ci. Здесь промысловая смертность каждого вида определяется независимыми промысловыми усилиями каждого из двух типов промысла. Модель была применена для двух конкурирующих за пищу популяций Юго-Восточной Атлантики – сардины и анчоуса (P - биомасса анчоуса, R –биомасса сардины):
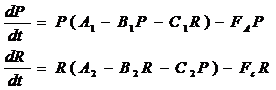 (3-6)
(3-6)
Эта модель при других обозначениях переменных и параметров практически совпадает с нашей моделью (2-14), но в данном случае авторы ставили задачу оценить параметры А, В, C (всего 6 параметров) для конкретной системы, и в качестве входной информации они использовали временные ряды P(t), R(t) и F(t), полученные методом одновидового ВПА. Входными данными для ВПА является возрастной состав уловов за ряд лет. Искомые параметры продукционной модели (3-6) определялись регрессионным методом при условии, что система находится в состоянии равновесия (т. е. при условии dP/dt= dR/dt =0). Такой метод оценки параметров не совсем корректен, поскольку, с одной стороны, методом ВПА определяется динамика запасов, что противоречит принятому условию равновесия системы, а с другой стороны, одновидовой метод ВПА не учитывает межвидовых отношений и потому может давать смещенные оценки запасов относительно реальной ситуации, которая должна описываться многовидовой моделью. Кроме того, если в распоряжении исследователей есть информация о возрастном составе уловов популяций, то на первый взгляд нет необходимости использовать продукционные модели для описания динамики популяций. Тем не менее, эти модели могут помочь оценить качество того или иного управления промыслом и выбрать наиболее близкое к оптимальному по выбранному критерию.
Оценив таким образом параметры модели, авторы построили область допустимого управления на плоскости {FA, FC } и показали, что на этой плоскости существуют подобласти, где один или другой вид могут исчезнуть, но существует и область, в которой обе популяции могут устойчиво существовать.
Итак, в обоих случаях (модели 3-5 и 3-6) авторы фактически ограничились рассмотрением стационарных задач, т. е. равновесия системы запас – промысел.
К. Сулливан (Sullivan, 1991) разработал продукционную модель трехвидового сообщества Балтийского моря, которая включала одного хищника-треску, два вида жертв (салаку и шпрот) и каннибализм трески. Входной информацией для продукционной модели являются оценки численности разных видов, которые были получены не по одновидовому ВПА, как в модели Поупа и Харриса, а с помощью многовидового размерного анализа (Pope, 1980). Этот анализ представляет собой расширение размерного одновидового анализа Джоунса (Jones, 1981), который сам является моделью равновесного состояния популяции. Для реализации же этого анализа понадобилась дополнительная информация по питанию хищника. Предполагалось, что рацион хищника постоянен и в нем постоянна доля рыбной пищи. Уровень питания и распределение коэффициента смертности от хищников M2 между видами жертв определялись из годового рациона, состав пищи и размерное предпочтение трески взяты из материалов рабочей группы по многовидовой оценке запасов Балтики (ICES, 1982). Функция Урсина (Ursin, 1973) описывает размерное предпочтение (в форме логнормального распределения, см. гл. 5), при этом коэффициент пищевой предпочтительности разных видов предполагается равным единице, т. е. хищничество считается зависимым только от размеров пищевых организмов.
В продукционной модели сообщества Балтийского моря рассмотрены два типа промысла – пелагический для салаки и шпрота и донный – для трески. Состояние равновесия для каждого нового значения F определяется итерационно, причем одновременно проводятся итерации нескольких процессов:
1) при новых значениях F меняются вылов и биомасса запаса каждого вида и
2) уровень хищничества и его влияние на молодь трески и на других жертв меняются при изменении относительных размеров трески и её распределения по размерам.
Параметры модели оцениваются с помощью уравнений многофакторной регрессии. После оценки параметров уравнения для состояния равновесия системы имеют следующий вид:
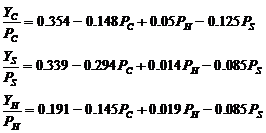
Индексы C, H, S при P (обозначение биомассы равновесного состояния популяции) и Y (вылов) соответствуют треске (cod), сельди (herring) и шпроту (sprat) соответственно. Расчеты показали, что в первом уравнении два последних слагаемых практически компенсируют друг друга, так как в среднем биомасса сельди в два с половиной раза выше биомассы шпрота. Потому запас балтийской трески хорошо описывается одновидовой моделью Шефера.
В уравнениях для сельди и шпрота проявляются сильные взаимодействия как с хищником, так и между видами жертв. Коэффициент при Pc во втором уравнении представляет собой долю биомассы шпрота (или сельди – в третьем уравнении), съеденной единицей биомассы трески, и этот коэффициент для сельди в два раза меньше, чем в уравнении для шпрота, это означает, что шпрот имеет более подходящие для трески размеры, чем сельдь. Уровень хищничества по отношению к сельди и шпроту примерно тот же – но поскольку биомасса сельди примерно в два раза выше, то шпрот можно считать более предпочтительным для трески.
Коэффициент при биомассе шпрота Ps в уравнении для сельди характеризует косвенное взаимодействие между сельдью и шпротом – через питание хищника. При высоком уровне интенсивности промысла трески ее средний размер уменьшается, и шпрот становится более пригодным для трески, при этом уменьшается влияние трески на сельдь. И наоборот – при уменьшении интенсивности промысла трески средний размер хищника увеличивается, а более крупная треска съедает больше сельди, и при высокой биомассе трески это может привести к резкому уменьшению биомассы сельди (коллапсу). Это тот случай, когда на первый взгляд непонятные знаки при членах уравнений, описывающих взаимоотношение видов, могут объясняться поведением хищника при питании, а именно его переключением на более пригодную пищу при изменении размерного состава популяции хищника.
Трехвидовая продукционная модель К. Сулливана (1991) тоже является равновесной моделью.
3.3 Многовидовая динамическая продукционная модель
В общем случае модель многовидового сообщества, состоящего из n видов, представляет собой, как правило, систему дифференциальных или конечно-разностных уравнений, зависящих от некоторых параметров. В простейшей ситуации, когда изменения численности (или биомассы) каждого вида описываются одним обыкновенным дифференциальным уравнением, такая система может быть представлена в виде:
dPi/dt = Fi(P1,P2,..,Pn,ai1,ai2,..aim ) – Yi; i=1,2,…, n (3-7)
где Pi(t) – обилие запаса i-го вида;
ail – коэффициенты i-го уравнения (l = 1, ....т); которые считаем постоянными величинами;
m – количество постоянных параметров каждого уравнения системы;
Fi – некоторые нелинейные функции относительно переменных P1,P2,..,Pn;
Yi – мгновенные уловы i - го вида (уловы, взятые в единицу времени).
Такая модель является динамической (или нестационарной), поскольку рассматривает изменение численности или биомассы во времени и поскольку, как будет показано ниже, предлагаемый метод оценки параметров модели учитывает изменчивость запасов.
Стационарные состояния системы (3-7) отвечают условиям равновесного промысла всех эксплуатируемых видов сообщества. В этом случае уравнения модели имеют вид Fi()=Yi, и метод определения параметров ail модели по промысловым данным принципиально не отличается от описанного выше для равновесной модели Шефера. Однако больший интерес представляют стационарные состояния системы (3-7), параметры которой определены с учетом динамики всех взаимодействующих запасов. Всюду в дальнейшем, где речь идет о стационарных состояниях системы (см. также гл. 4), они понимаются именно в таком смысле.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
