
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Прогностическая способность модели оценивается по степени соответствия этих рядов в течение прогнозного периода. Для варианта, когда параметры оцениваются для поколений Y=1984-2000 гг., прогнозным периодом модели считаем интервал, начинающийся с Y=2004 г., т. е. сопоставление рядов возможно на четырехлетнем интервале Y =2004-2007 гг. На этом интервале коэффициент корреляции между рядами R(Y) и Rmod(Y) равен 0,70 для IUU-Росс и 0.75 для IUU-Норв.
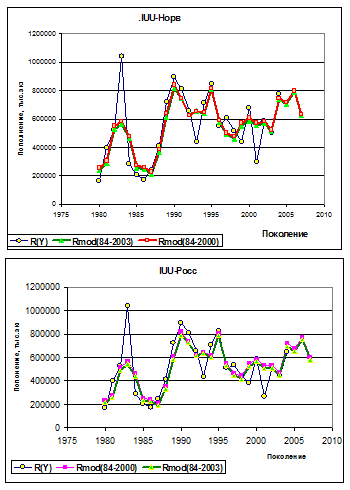
Рисунок 7.4. Результаты параметризации модели (7-3), полученные на основе двух прогонов XSA (ICES, 2008): при IUU-Норв (верхний рисунок) и IUU-Росс (нижний рисунок). Оценки пополнения R(Y), полученные по прогону XSA, сопоставлены с модельными Rmod при двух интервалах параметризации модели (Y=1984-2000 гг. и Y=1984-2003 гг.)
Таблица 7.2
Прогностические значения величины пополнения (млн. экз.), полученные по модели (7-3) для двух прогонов XSA и разных интервалов лет, используемых при параметризации модели пополнения трески
Поколение Y | 2004 | 2005 | 2006 | 2007 |
IUU-Росс | ||||
1984-2000 | 716 | 674 | 772 | 600 |
1984-2003 | 696 | 651 | 753 | 579 |
IUU-Норв | ||||
1984-2000 | 743 | 715 | 798 | 633 |
1984-2003 | 738 | 705 | 790 | 624 |
На двух последних заседаниях Арктической группы ИКЕС (ICES 2007; 2008) оценка запаса трески проводилась не только по методу XSA, но и по модели TISVPA (Triple Instantaneous Separable VPA), которая является развитием сепарабельной модели ISVPA (Vasilyev, 2005; 2006). Отличие этой версии сепарабельной модели от предыдущей (ISVPA) в том, что коэффициенты промысловой смертности представляются в виде произведения не двух параметров f(year)*s(age), а трех: f(year)*s(age)*g(cohort), причем третий множитель описывает зависимость смертности от поколения. Это позволяет описывать ситуацию, при которой разные поколения могут по - разному взаимодействовать с промыслом.
Дело в том, что модель XSA имеет ряд недостатков, из которых основным является то, что ее решение получается в результате сходимости некоторой итерационной процедуры, а не путем нахождения экстремума целевой функции. Эта процедура сходится не всегда, но если и сходится, то нет уверенности, что она сходится к единственному верному решению. Модели же класса ISVPA не имеют этих недостатков и, что очень важно, способны работать с сильно зашумленными входными данными.
Таблица 7.3
Результаты параметризации модели пополнения, полученные на основе прогона TISVPA по данным IUU-Росс. Значения индекса N(Y) рассчитаны по аномалиям температуры относительно линейного тренда
Поколения, выбранные для оценки параметров | 1984-2000 | 1984-2003 |
Количество точек / степеней свободы | 17/13 | 20/16 |
A | 4,53 | 4,60 |
Can | 1,72*10-6 | 1,69*10-6 |
ny | 0,0522 | 0,.0564 |
r2 | 0,75 | 0,65 |
Оценки и нерестового запаса трески, и ее пополнения, полученные по модели TISVPA (Васильев и Булгакова, 2007; Vasilyev and Bulgakova, 2007) несколько отличаются от результатов XSA, поэтому по новым данным тоже была проведена параметризация модели пополнения.
Видим (табл.7.3), что модель пополнения, параметры которой найдены по результатам TISVPA, описывает 65-75% изменчивости пополнения трески, что близко к оценкам, полученным по результатам XSA (табл. 7.1).
Таким образом, можно утверждать, что успех пополнения арктической трески зависит от величины родительского стада и от его структуры, от уровня каннибализма и от условий внешней среды. Отсутствие отчетливой связи между биомассой нерестового запаса и численностью нарождающегося от него поколения не означает, что этой зависимости нет, просто она завуалирована влиянием других факторов.
Процессы формирования пополнения арктической трески давно интересовали исследователей, и чаще всего авторы ограничивались статистическими методами, пытаясь нащупать факторы, влияющие на эти процессы. В. Бойцов и Р. Мельянцев (1994) исследовали влияние таких факторов как популяционная плодовитость, процент и численность самок, численность самок не старше 14 лет и в возрасте 9-14, средневзвешенный возраст нерестовой части стада, а также температура воды в разные периоды года и на разных станциях основной ветви Мурманского течения, в том числе средняя и минимальная температура в период зимовки молоди. Для периода после 1979 г. авторы установили фактор, частотная характеристика которого близка к частотной характеристике пополнения трески в возрасте трех лет – это средняя температура во время первой зимовки (март-май) в южной части Баренцева моря – на разрезах 6, 8 и 10 в слое 50-200м. (r =0.810).
Нильсен и др. (Nilssen et al. 1994) применили уравнение множественной регрессии численности пополнения 0-группы по трем факторам: температуре на Кольском разрезе1,[10]биомассе нерестового запаса и суммарной численности молоди в возрасте трех и четырех лет (эта величина выбрана авторами в качестве индекса каннибализма). Полученная зависимость объясняет около 46% изменчивости пополнения для периода 1946-1988 гг., при этом авторы подтвердили существование статистически значимой регрессионной зависимости пополнения от запаса.
с соавторами (Stiansen et al. 2002) разработали базу данных, включающую около 70 временных рядов климатических переменных и характеристик основных промысловых популяций. с соавторами (Stiansen et al. 2003) построили уравнения множественной регрессии для описания и прогнозирования численности сеголетков трески, используя логарифм численности сеголеток R(0+) в качестве зависимой переменной, а в качестве независимых факторов взяв зимний индекс NAO2[11]и SSB – оба фактора введены в модель с заблаговременностью два года. При этом получен довольно высокий коэффициент корреляции, равный 0,8. Сомнение вызывает биологический смысл таких задержек, авторы трактуют их как время, необходимое для развития планктонных популяций, представляющих корм для молоди трески, и для их переноса в район Баренцева моря.
В нашей модели (7-3) введены два дополнительных (помимо каннибализма) фактора. Первый из них – популяционная плодовитость. Этот показатель, как показано выше, рассчитывался приблизительно, т. е. при некоторых допущениях. Уточнение его можно провести, заменив его на альтернативный энергетический показатель, например, на величину общей липидной энергии ОЛЭ (Marshall et al., 2000). Упомянутые авторы провели линейный многофакторный анализ для исследования зависимости величины пополнения трески от ОЛЭ в печени половозрелых самок и температуры воды на Кольском разрезе (в период с апреля по июль) и показали, что такая модель описывает до 43% изменчивости пополнения за достаточно длинный период – около 50 лет. Плотностную зависимость эти авторы не рассматривали.
Структура нерестового запаса характеризуется не только его возрастным составом, но и таким показателем, как доля повторно нерестующих особей в стаде. Икра, полученная от повторно нерестующих самок, имеет меньшую смертность, чем выметанная впервые нерестующими (Marshall et al., 2000; Tretyak, 2003). Примерно после 1980 г. изменилась качественная структура нерестового стада трески, в те же годы резко уменьшилась доля поздно созревающих особей (Tretyak, 2003). А это означает, что ухудшилось качество половых продуктов трески, а следовательно и выживаемость поколения до трех лет.
Включение в модель второго экосистемного фактора – индекса установившегося притока Атлантических вод NY(Y) позволило существенно увеличить корреляционное отношение, т. е. более точно описать динамику пополнения.
По нашей модели пополнения трески построена двухмерная зависимость пополнения от SSB и индекса N(Y) (рис.7.5). Индекс m(Y), равный отношению популяционной плодовитости к биомассе нерестового запаса, который линейно входит в зависимость (7-2), при этом предполагался равным среднемноголетней величине, рассчитанной по ретроспективным оценкам. На рис.7.6 показаны предельные значения функции Rmod от SSB, т. е. верхняя предельная функция соответствует N(Y)=12 и максимальному значению m(Y), а нижняя – N(Y)=0 и минимальному значению индекса m(Y). Понятно, что реальная изменчивость пополнения может быть еще выше, поскольку здесь не были учтены случайные факторы.
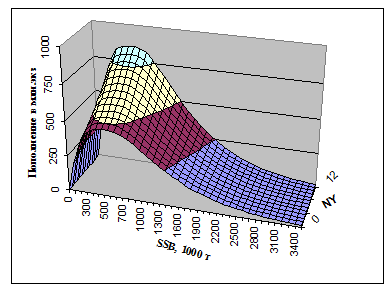
Рис.7.5. Пополнение как функция двух переменных SSB и NY согласно модели (7.2) при среднемноголетнем значении индекса m(Y).
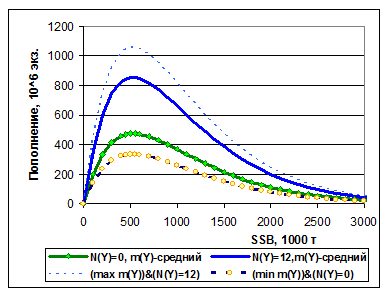
Рисунок 7.6. Предельные значения детерминированной функции R(SSB) согласно модели (7-3). Сплошные линии соответствуют среднему значению относительного индекса популяционной плодовитости m(Y), пунктирные – максимальному значению m(Y) и N(Y)=12 (верхняя) и минимальному значению m(Y) и N(Y)=0 (нижняя).
Попытка распространить уравнение (7-3) на весь период, начиная с 1946 г., не удалась – модель плохо описывает динамику пополнения, особенно плохо аппроксимируются пики годовых классов 1963-1964 и 1970 гг. Таким образом, с точки зрения температурных характеристик и показателя притока Атлантических вод мощность этих поколений объяснить не удается. Не исключено, что для первого периода следует отдельно искать параметры зависимости запас-пополнение или даже включать в эту зависимость другие факторы среды.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
