
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Год промысла | Возраст рыб в улове | ||||
3 | 4 | 5 | …. | n | |
t | Ct,3 | Ct,4 | Ct,5 | …. | Ct, n |
t+1 | Ct+1,3 | Ct+1,4 | Ct+1,5 | …. | Ct+1,n |
t+2 | Ct+2,3 | Ct+2,4 | Ct+2,5 | …. | Ct+2,n |
… | … | … | … | …. | …. |
… | … | … | … | …. | …. |
t+k | Ct+k,3 | Ct+k,4 | Ct+k,5 | …. | Ct+k, n |
Очевидно, что по любой диагонали таблицы стоят численности рыб, взятых промыслом в разные годы из одного и того же поколения. При предположении, что в течение временного интервала в один год для каждого возраста a параметры смертности не меняются, можно записать ряд уравнений, используемых для оценки численности запаса по ВПА. Расчет ведется для каждого поколения отдельно, и для упрощения записи в (5-1) опущен индекс года:
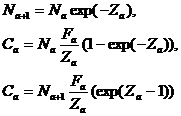 . (5-1)
. (5-1)
Второе уравнение системы (5-1) связывает величину годового улова, взятого из а-ой возрастной группы (Ca), с численностью той же группы в начале года, а третье уравнение получается при подстановке Na+1 из первого уравнения во второе и отражает зависимость величины того же улова от численности той же возрастной группы в конце года, которая равна численности следующей возрастной группы в начале следующего года.
Одновидовой ВПА требует наличия следующей входной информации:
· возрастной состав уловов за ряд лет;
· параметр естественной смертности Ma, t – в общем случае можно учитывать его зависимость от возраста и от времени, но часто принимается допущение, что этот параметр имеет одно и то же значение для всей промысловой части популяции;
· Fst – коэффициент промысловой смертности самой старшей возрастной группы выбранного поколения, представленной в уловах;
· wa, t – средний вес особи возраста a в год промысла t;
· темп полового созревания.
Последние два параметры нужны для расчетов биомассы запаса. Расчеты проводятся от старших возрастов к младшим в следующем порядке.
1. Сначала для выбранного поколения оценивается численность самой старшей группы с индексом n в начале года с помощью второго уравнения (5-1):
![]() ;
;
2. для предыдущей (n-1)-ой возрастной группы по третьему уравнению (5-1) методом последовательных приближений определяется величина Fn-1 (поскольку этот параметр входит в уравнение и в линейном виде и в показатель экспоненты);
3. по первому уравнению (5-1) определяется численность Nn-1 в начале года;
4. последовательно рассчитываются коэффициенты промысловой смертности и численности для более младших возрастных групп. Затем переходим к следующему поколению.
В результате расчетов получаются матрицы коэффициентов промысловой смертности и численности для всех лет и возрастов, структура их та же, что приведена в табл. 5-1. Численность промыслового запаса по годам промысла рассчитывается суммированием численности возрастных групп по строкам, биомасса запаса получается умножением численности возрастных групп на wa, t. Зная темп полового созревания, можно оценивать биомассу нерестового запаса. Таким образом, ВПА дает возможность получить ретроспективные оценки промыслового и нерестового запасов.
Наиболее сложным моментом метода ВПА является определение стартовых значений промысловой смертности. Разработаны несколько процедур их оценки, которые названы настройкой ВПА. Наиболее часто употребляемые методы настройки описаны Дж. Поупом и Дж. Шепардом (Pope and Shepherd, 1985). Для выполнения этих процедур необходима дополнительная информация, представленная в виде временных рядов дифференцированных по возрастам индексов запаса, которые иногда удается получить по учетным съемкам или по данным улова на единицу промыслового усилия (так называемые CPUE).
К когортным методам относится и метод размерных когорт Джоунса (Jones, 1981), который применяют для оценки запасов в тех случаях, когда отсутствуют данные по возрастному составу уловов, но доступны данные по размерному составу для нескольких лет. Необходимой входной информацией для этого метода являются также параметры уравнения роста Берталанфи (зависимости размера особей от возраста). Нами разработан аналог метода Джоунса для другого уравнения роста –Паркера-Ларкина (Parker and Larkin,1959), и по этим двум методам проведены расчеты запаса для капской ставриды (Babayan and Bulgakova, 1983; Булгакова и Бабаян, 1985). Несмотря на то, что этот метод является когортным, практически он оценивает величину запаса в состоянии равновесия.
5.2. Многовидовая когортная модель MSVPA
Одним из необходимых параметров при оценке запасов является коэффициент естественной смертности M, который оказывает значительное влияние на результаты. Но этот параметр является и одним из наиболее сложно определяемых параметров популяции. Причина этой сложности лежит в аддитивном характере коэффициентов промысловой и естественной смертности – даже если оценен параметр Z, проблема состоит в разделении его на составляющие. Если, например, оценка параметра M завышена, то в расчетах ВПА параметр F получится заниженным, в результате будет завышена оценка запаса.
Существует несколько методов оценки коэффициента естественной смертности, при этом чаще всего оценивается его средняя величина для промысловых возрастов популяции. Обзор существующих методов оценки M приводится в работах (1976), У. Рикера (1983), с соавторами (1984). В отечественной литературе существуют также работы, в которых делаются попытки оценить зависимость естественной смертности от возраста (Тюрин, 1972; Зыков и Слепокуров, 1982; Блинов, 1979). получает зависимость M(a) (точнее, естественной убыли) для мало облавливаемых популяций и на многих примерах показывает, что минимум этой кривой приходится на возраст полового созревания. Работа эта вызвала большой интерес исследователей, но в ней есть и слабые места (Печников, 1989). и строят зависимость M(a), заранее произвольно придав ей форму квадратичной параболы. (1979) построил эту зависимость для частных случаев: для камбалы Северного моря, полагая известным и постоянным коэффициент промысловой смертности, и для озерных популяций рыб по данным, полученным после тотальной химической обработки озер.
Нами был разработан (Булгакова и Ефимов, 1982; Булгакова, 1985) метод, позволяющий включить зависимость смертности от возраста M(a) в расчет возможного улова, что расширило возможности классической модели промысловой популяции Р. Бивертона и С. Холта (1969).
Оценки коэффициентов естественной смертности (или выживаемости) можно оценивать и с помощью мечения особей. При этом следует учитывать и "издержки" мечения, а именно вероятность потери меток и дополнительную смертность от мечения. Этим вопросам посвящены работы В. Андреева, Т. Булгаковой и Ф. Челнокова (1976, 1978) в применении к популяциям морских котиков Командорских островов и о. Тюленьего.
Основное положение многовидового когортного анализа, обозначаемого MSVPA (MultiSpecies Virtual Population Analysis), заключается в том, что для каждого вида сообщества коэффициент естественной смертности подразделяется на две компоненты ![]() , где i −индекс вида, а − индекс возрастной группы. Первое слагаемое представляет собой коэффициент смертности от хищников особей возрастной группы а вида i, а второе слагаемое – коэффициент естественной смертности той же возрастной группы от всех других причин, который назван остаточной естественной смертностью.
, где i −индекс вида, а − индекс возрастной группы. Первое слагаемое представляет собой коэффициент смертности от хищников особей возрастной группы а вида i, а второе слагаемое – коэффициент естественной смертности той же возрастной группы от всех других причин, который назван остаточной естественной смертностью.
Н. Даан (Daan,1973) впервые поставил под сомнение гипотезу о постоянстве во времени параметра смертности популяции от хищничества M2. Этот параметр может зависеть не только от возраста особей, а также от численности и возрастного состава популяций хищников. Межгодовая изменчивость численности разных кормовых объектов и их потребителей также приводит к изменчивости параметра М2. Таким образом, одной из целей разработки модели MSVPA было стремление снизить неопределенность в используемых значениях коэффициентов естественной смертности и учесть межвозрастную и межгодовую изменчивость этого параметра.
Большое значение для развития многовидового моделирования имела работа К. Андерсена и Е. Урсина (Andersen & Ursin, 1977), которые предложили очень детальную математическую модель экосистемы, включающую разные трофические уровни – от планктона до промысловых рыб; модель также описывала циркуляцию биогенов, стратификацию вод, миграцию животных и пр. Математический аппарат этой модели – дифференциальные уравнения, как системы обыкновенных дифференциальных уравнений, так и в частных производных при описании неоднородностей процессов, происходящих в океане. Ценность этой модели в том, что ее можно использовать как справочник при моделировании отдельных процессов в экосистеме. Но большое количество моделируемых процессов и множество неизвестных параметров не позволяют использовать ее целиком в качестве модели экосистемы. Хотя отдельные блоки этой модели были применены и развиты далее при моделировании, например, сообщества Балтийского моря (Horbowy, 1989; 1993).
Стартовой точкой для развития именно когортной модели MSVPA принято считать доклады Дж. Поупа, а также Т. Хелгасона и Х. Гисласона, представленные на Ежегодную научную конференцию ИКЕС в 1979 г. (Роре, 1979; Helgason and Gislason, 1979). Подробное описание алгоритма MSVPA приведено в работе (Gislason and Helgason, 1985), а первая версия компьютерной программы предложена П. Спарре (Sparre, 1984). Затем программа много раз усовершенствовалась группой датских ученых (Gislason and Sparre, 1987; Sparre, 1991).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
