
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
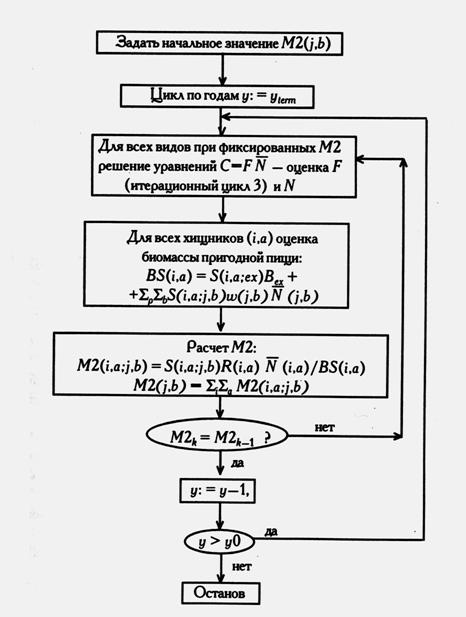
Рисунок 5.3. Итерационные циклы 2 и 3 вычислительной процедуры MSVPA при уже оцененных значениях S(), k – номер итерации решения уравнений относительно M2() (по Sparre, 1991).
Таким образом, в модели MSVPA, для которой разработана компьютерная программа, функция пригодности пищи для каждого хищника рассчитывается внутри модели. Возможен и иной подход, при котором эта функции описывается в том виде, в котором она была предложена К. Андерсеном и Е. Урсином (1977), а точнее, несколько раньше Е. Урсин (Ursin, 1973) построил эту зависимость по конкретным данным по питанию трески и камбалы-лиманды. Он показал, что для рыб Северного моря для каждого размера (массы) потребителя есть наиболее предпочитаемый размер жертв, и что особи жертв, имеющие массу в 2 раза больший и в 2 раза меньший предпочитаемого размера, одинаково часто встречаются в желудках этого хищника. Таким образом, функция предпочтительности, у которой по оси абсцисс отложен логарифм отношения массы хищника к массе жертвы, должна быть симметрична относительно точки η(i) – натурального логарифма оптимального отношения массы хищника к массе жертвы. Функция пригодности, согласно Е. Урсину (1973), имеет вид:
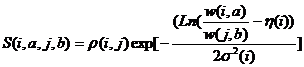 (5-11)
(5-11)
Первый множитель в уравнении (5-11) – это коэффициент уязвимости j-го вида как пищи для i-го вида хищника, показывающий, например, какая доля особей j-го вида расположена в тех же слоях воды, что и хищник. Если некоторый вид жертв, по размерам подходящий для данного хищника, не встречается в его желудках вообще, то в этом случае ρ(i, j) = 0. Второй множитель представляет собой функцию размерной предпочтительности пищи, которая характеризуется двумя параметрами – оптимальным отношением массы хищника к массе жертвы η(i)для i-ого вида и его стандартным отклонением σ(i). Если эти параметры оценить заранее (вне модели), то с одной стороны, упростится алгоритм (не нужен будет цикл итераций поиска функции S()), а с другой стороны, функция предпочтительности будет оцениваться сравнительно просто для каждого интервала времени и сможет учитывать переменный темп роста особей разных видов. Такой подход был предложен Т. Хелгасоном и Х. Гисласоном (Helgason and Gislason, 1979) и применен в нескольких работах, в том числе в нашей модели сообщества экосистемы шельфа Намибии (Bulgakova and Kitain, 1985), в которой трофические взаимоотношения разных видов характеризуются только отношением размеров особей потребителей и жертв (Булгакова, 1987).
Упрощенным вариантом MSVPA является одновидовая модель с учетом каннибализма, предложенная испанскими учеными (Lleonart, Salat, Macpherson, 1985) для популяции капского хека. Понятно, что расчеты в этом случае менее громоздки, и легче анализировать результаты. Авторы пришли к выводу, что традиционная одновидовая модель ВПА примерно в полтора раза занижает оценки смертности и биомассы для младших возрастов хека.
5.2.4. Прогностический блок многовидовой модели
Программный блок ретроспективного многовидового анализа, алгоритмы которого описаны выше, формирует основной выходной файл, где приводятся оценки разных популяционных параметров, перечисленных в предыдущем параграфе, а также два вспомогательных файла, которые затем служат входными файлами для прогностического блока. В этих файлах в специальных форматах записаны результаты расчетов ретроспективного блока, в том числе средние многолетние оценки веса особи разных видов по возрастным группам, средние рационы, функция пригодности разных категорий пищи, а также оценки численности для последнего (терминального) года ретроспективных расчетов. Еще в один файл пользователь записывает выбранные им условия проведения прогнозных расчетов, например, количество прогнозных лет, величину пополнения для каждого из видов, масштабные коэффициенты, которые позволяют варьировать коэффициенты промысловой смертности видов при прогнозировании и др.
Прогностические расчеты ведутся в основном по тем же формулам, что и при одновидовом прогнозировании, за исключением смертности от хищничества, которая оценивается по тем же формулам, что и в ретроспективном блоке MSVPA. В данном случае функция пригодности пищи берется та же, что получена в результате итерационных расчетов после усреднения коэффициентов S() по годам, для которых существует информация по составу желудков. Величина пополнения каждого вида принимается постоянной на весь прогнозный период. В программе заложена возможность статистических расчетов, но для использования этой возможности требуется мощная оперативная система типа UNIX и статистический пакет SAS.
В настоящее время модель MSVPA получила широкое распространение в практике оценки запасов – помимо Северного моря, она используется для сообщества рыб Балтийского моря (Sparholt, 1991), Баренцева моря (Bulgakova et al., 1995c; Коржев и Долгов, 1999; Многовидовой анализ…, 2001), банки Джорджес (Tsou and Collie, 1998).
Глава 6. Исследование функционирования промыслового
сообщества Баренцева моря
Модель MSVPA впервые была применена к экосистеме Северного моря и в течение нескольких лет неоднократно модифицировалась (Gislason and Sparre, 1987; Sparre,1991; Pope, 1991). Работе с этой моделью была посвящена специальная рабочая группа ИКЕС по многовидовым оценкам запасов, потом была организована рабочая группа по многовидовым оценкам запасов Балтики.
При работе с сообществом Баренцева моря за основу нами выбрана версия модели, разработанная для Балтийского моря, позволяющая задавать переменный по годам темп роста массы особей в уловах и в запасе (в море). Эта версия была нами модифицирована так, чтобы можно было задавать переменные по кварталам коэффициенты M1, что важно для более адекватного описания сезонной динамики мойвы Баренцева моря, характеризующейся чрезвычайно высокой смертностью после нереста и сезонным характером промысла (Bulgakova et al., 1995с). С другой стороны, пришлось ввести в программу возможность задавать переменные по годам и кварталам рационы хищника, поскольку было показано (Булгакова и др., 1995; Bulgakova et al., 1995a), что рационы трески Баренцева моря сильно варьируют во времени, что по-видимому вообще характерно для хищников бореальных сообществ, в которых сильно варьируют и темп роста и состав пищи хищников (Mehl and Sunnanå, 1991).
6.1. Структура сообщества Баренцева моря
В состав моделируемого сообщества включены виды, составляющие "ближайшее окружение" северо-восточной арктической трески (Gadus morhua L.) – вида, продукционные возможности которого во многом определяют экономику прибрежных государств. В сообщество, помимо трески, вошли мойва (Mallotus villosus), северная креветка (Pandalus borealis) и атлантическо-скандинавская сельдь (Clupea harengus L.) – эти три вида представляют собой основные компоненты питания трески. Кроме того, моделируется и каннибализм трески, поскольку он сильно влияет на формирование ее пополнения.
Возрастная структура популяций моделируемого сообщества приведена в табл. 6.1. Ретроспективные расчеты проводились для интервала лет с 1980 по 1996 гг. с шагом в один квартал.
Таблица 6.1
Вариант структуры моделируемого сообщества Баренцева моря
Вид | Возрастные группы в модели | Возраст хищника | Возраст жертвы |
Треска | 0+- 15+ | 1 - 15+ | 0+- 4 |
Мойва | 0+ - 6 | - | 0+- 6 |
Креветка | 0+ - 6 | - | 0+- 6 |
Сельдь | 0+ - 12+ | - | 0+- 4 |
6.2. Подготовка входной информации
Для работы с моделью MSVPA требуется большой объем разнородной информации. Кроме данных по каждому виду, традиционно используемых при моделировании одной популяции, необходимы данные для описания взаимодействий между видами. Прежде всего, это информация по составу желудков. Для ее получения требуется ежегодно (а для проведения расчетов на квартальной основе – ежеквартально) проводить массовый сбор и весьма трудоемкий анализ содержимого желудков. При этом необходимо, чтобы пробы по питанию хищника разных возрастных групп были достаточно представительны, а также взяты равномерно по всему ареалу вида. Поэтому практически значимое использование модели MSVPA стало возможно лишь после накопления результатов многолетних работ по анализу содержимого желудков. Например, в начальный период работы над моделью MSVPA сообщества Северного моря, с 1984 г. по 1986 г., использовались данные по составу желудков хищников только за один 1981 г., но эти данные были собраны учеными разных стран по специально разработанной международной программе сбора материалов, так называемой «Программы желудка 1981 года». Постепенное накопление материала позволило включать данные и за последующие годы, а в 1991 г. снова была проведена международная съемка в Северном море, позволившая собрать представительный материал по питанию разных хищных рыб.
Советские ученые начали собирать материал по питанию трески Баренцева моря еще в 20-е годы прошлого столетия. В послевоенные годы такие материалы собирались в ПИНРО и в Мурманском морском биологическом институте уже более или менее регулярно. Подробный анализ исследований по питанию трески приведен в монографии ПИНРО «Треска Баренцева моря» (1996) и в работах российских и норвежских ученых (Новикова, 1965; Пономаренко, 1974; Тарвердиева и Ярагина 1989;1996; Богстад и Мель, 1992; Mehl and Yaragina, 1992 и др., Долгов, 1999; Пономаренко и Ярагина, 2003; Dolgov et al, 2008).
Развитию многовидового моделирования для Баренцева моря способствовало создание совместной Российско-Норвежской компьютерной базы данных (БД) по питанию хищников. Работа над её созданием была начата в 1988 г. совместно учеными России и Норвегии: были разработаны единая структура базы данных, кодификаторы гидробионтов, формы для сбора информации и т. д. (Третьяк и др.,1990). Затем было разработано специальное математическое и программное обеспечение для ведения этой базы данных.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
