
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
![]() (1-19)
(1-19)
Функции Ki должны удовлетворять ряду ограничений, и в зависимости от системы ограничений модель описывает систему взаимодействующих видов по типу хищник-жертва, конкуренция либо симбиоз. На плоскости {x1,x2} авторы строят линии K1=0 и K2=0, соответствующие горизонтальной и вертикальной изоклинам. Если они пересекаются в положительной области, то их точка пересечения соответствует состоянию равновесия.
А. Рисайо и К. Джонс (Rescigno and Jones, 1972) обобщают модель (1-19) для случая трофической цепи разной длины. Авторы делают ряд правдоподобных гипотез относительно возможных изменений численностей, из этих гипотез выводятся свойства функций Ki . Для n=3 показано, что может существовать только одна точка равновесия, при которой все виды сосуществуют, а решение может быть периодическим. (В отличие от простой модели Вольтерра, для которой для нечетного числа видов точки равновесия нет). Тот же вывод распространяется и на сообщество, представляющее собой трофическую цепь n видов.
Дж. Вандермеер (Vandermeer, 1973) графически показывает, как при разном взаимном расположении изоклин и разной их форме получаются разные типы взаимоотношений видов и разные типы особых точек.
1.2.6.Устойчивость элементарной модели с лимитирующими факторами
В предыдущих параграфах рассматривались математические модели, в которых вид функций в правых частях уравнений не меняется со временем. (1966, 1973) разработал новые принципы построения моделей биогеоценозов, используя принцип лимитирующих факторов Либиха – так называемые L-системы.
В общем случае моделируется система видов (популяций), обитающих на одной территории. Каждый k-ый вид имеет в своем распоряжении несколько типов корма (в том числе к кормам могут быть отнесены некоторые популяции экосистемы), а каждый тип корма состоит из ряда компонент. Если состав каждого корма постоянен, то его количество можно представить в виде вектора 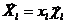 , где вектор
, где вектор ![]() характеризует состав корма, некоторые из составляющих которого могут быть равны нулю.
характеризует состав корма, некоторые из составляющих которого могут быть равны нулю.
В сообществе происходят различные процессы, характеризующиеся интенсивностями Pj и влияющие на изменения численности популяций. Например, сообщество, состоящее из двух популяций, хищника и жертв, можно описать системой следующих дифференциальных уравнений (Полетаев, 1973):
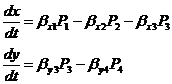 (1-20)
(1-20)
Здесь β – положительные константы, P1– интенсивность размножения жертвы, P2 и P4 – естественная смертность видов жертв и хищников соответственно, P3 – интенсивность потребления жертв хищниками и пропорциональная ей интенсивность размножение хищников. В каждом процессе по аналогии с химическими процессами потребляются одни вещества и выдаются вовне другие. Если в единицу времени для протекания j-го процесса (на единицу его интенсивности) необходимо потреблять несколько веществ с индексом i, в количестве αij , а поток или запас этих веществ равен ωij, то интенсивность j -го процесса определяется по формуле
![]() .
.
Эта формула и представляет собой принцип Либиха: именно вещество с индексом i, при котором при данного состоянии системы отношение в скобках минимально, и является лимитирующим фактором для j-го процесса, т. е. количество именно этого вещества определяет интенсивность протекания процесса. Понятно, что как величины ωij, так и интенсивности протекающих процессов Pj переменны во времени и зависят от численностей популяций x и y. При изменении состояния системы {x, y} могут смениться лимитирующие факторы – и так для каждого из протекающих в экосистеме процессов.
Для системы уравнений (1-20) интенсивности процессов можно представить в следующем виде:
![]() . (1-21)
. (1-21)
Содержательно (1973) трактует эти формулы следующим образом. Размножение жертв определяется как численностью самой популяции, так и некоторым внешним фактором E, например, потоком света для растений или площадью нерестилищ для рыб. Здесь принято, что E=const. Интенсивности процессов естественной смертности (P2 и P4) определяются только численностью соответствующей популяции, потому знак минимума перед ними опущен. Процесс потребления жертв и размножения хищников P3 лимитируется либо поголовьем хищников, либо количеством корма, потребление которого, как и В. Вольтерра, считаем пропорциональным числу встреч (произведением численностей), или внешним фактором F. Все параметры α и λ считаются постоянными величинами.
При заданных начальных условиях {x0,y0} система (1-20) имеет единственное решение. При решении системы уравнений следует подставить начальные условия в уравнения (1-21), а затем полученные значения Pj –в систему уравнений (1-20). Вся область решений разделяется на несколько областей, границы которых соответствуют переключению одного из лимитирующих факторов на другой, при этом изменяется вид функции Pj , т. е. и правой части системы (1-20). В зависимости от начальных условий и значений параметров решение системы (1-20) может иметь самый разный вид, в том числе разное количество точек равновесия и разные свойства этих точек (Полетаев, 1973). Показано, что модель Вольтерра является частным случаем системы (1-20).
В нашей работе (Булгакова, 1968) рассматривается элементарная ячейка системы с лимитирующими факторами и исследованы условия существования устойчивой точки равновесия такой системы. Пусть элементарная система состоит из одного вида-потребителя (численностью z) и одного типа корма (переменное количество корма x), который пополняется с постоянной скоростью C.
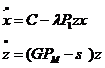 (1-22)
(1-22)
Потребитель развивает три активности – основной обмен (затраты на него равны постоянной величине b0), добыча корма P1 и размножение PM, затраты на две последние активности пропорциональны их интенсивности, b1P1 и bMPM соответственно.
Допустимые значения активностей потребителя ограничены следующими неравенствами, где первое является ограничением на физиологические возможности организмов развивать указанные выше активности, а второе описывает условие сохранения вещества или энергии:
![]() (1-23)
(1-23)
Здесь α – содержание критической компоненты в единице массы корма. Параметры α, C, G, s (коэффициент смертности) и φ– постоянные величины.
Область возможных значений активностей P1 и PM на плоскости {P1, PM} заштрихована на рис. 1.12, и если предположить, что потребитель будет стараться максимизировать активность размножения, то выбранные им активности будут соответствовать точке B, координаты которой определяются совместным решением уравнений (1-23), полученных после замены знаков неравенств на равенства:
![]() , (1-24)
, (1-24)
где 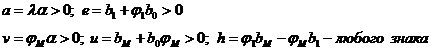 .
.
Из (1-24) видно, что допустимые значения количества корма ограничены неравенством x≥e/α.
Подставив значения ![]() из (1-24) в (1-22), получим систему уравнений
из (1-24) в (1-22), получим систему уравнений
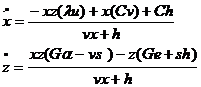 .
.
Исследование этой системы уравнений показало (Булгакова, 1968), что стационарная точка экосистемы существует в допустимой области при условии G-sφM>0, и эта точка устойчива при h>0. Ясно, что поскольку h=φ1bM - φMb1, с ростом затрат на единицу интенсивности добычи корма b1 устойчивость системы ослабляется (рис. 1.13).
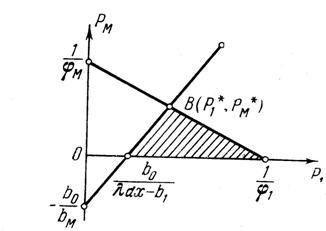
Рисунок 1.12. Область возможных значений интенсивностей двух процессов заштрихована. Точка В соответствует максимальной интенсивности размножения популяции
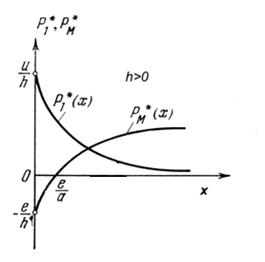
Рисунок 1.13. Зависимость интенсивностей добычи корма и размножения потребителя от количества корма x при h>0.
В исследованной выше простой модели рассматривается только один вид корма, содержащий один существенный компонент. Но даже один тип корма может содержать и два важных для развития потребителя компонента, в этом случае при той же системе уравнений (1-22) к ограничениям (1-23) должно быть добавлено еще одно, аналогичное второму, но с другими коэффициентами:
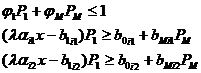 ,
,
В полученной системе возможны ситуации, когда один лимитирующий фактор при некотором значении x заменяется другим. Это значение обозначим как xкр– при значенияx xmin<x< xкр вид-потребитель ограничен первой компонентой, а при x>xкр лимитирующим фактором становится вторая компонента. Таким образом, область значений x разделена на две области.
Численные расчеты по такой модели показали, что даже если в одной из областей решение неустойчиво, а в другой устойчиво, переключение лимитирующих факторов может быть причиной асимптотической устойчивости общего решения.
Дальнейшие теоретические разработки методологии приведены в работе Ю. Гильдермана с соавторами (1970), на той же основе построена модель нижних трофических уровней черноморской экосистемы (Айзаттуллин, Хайлов, 1973).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
