
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
2. Проводится замена переменных yi =xi - xio , тогда {yi} –приведенная система, т. е. состояние равновесия системы (1-1) xi= xio соответствует тривиальному решению приведенной системы.
3. Рассматривается вопрос устойчивости решений приведенной системы относительно ее тривиального решения. Определяются знаки действительных частей корней характеристического уравнения линеаризованной системы уравнений. Для того чтобы корни алгебраического уравнения имели отрицательные действительные части, необходимо и достаточно, чтобы все определители Гурвица этого уравнения были положительны. Для характеристического уравнения вида
a0λn+a1 λn-1+a2 λn-2+…an=0
определители Гурвица записываются в следующем виде
Δ1=a1; Δ2=![]()
![]() ;….; Δn=
;….; Δn=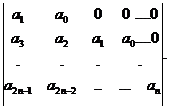 ,
,
где ai= 0 при i>n.
Проведем исследование этим методом модели Вольтерра (1-1) для n видов.
1. Точка равновесия системы (1-1) определяется из системы алгебраических уравнений, получаемой приравниванием нулю правых частей уравнений (1-1). Ищется решение этой системы {xio}, которое лежит в положительной области. Если в какой-то момент времени один из компонентов (звеньев) (i=i0) равен нулю, то это звено трофической цепи уже не может восстановиться, и все последующие звенья тоже будут исчезать, поскольку первая производная для них становится отрицательной. В результате останется система порядка (i0-1).
Определяем координаты равновесной точки {xio} из линейной алгебраической системы уравнений
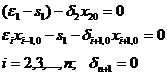
Определитель D этой системы равен
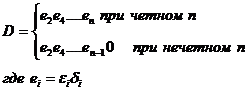
Система алгебраических уравнений имеет единственное решение, если ее определитель не равен нулю. Отсюда следует, что при нечетном n уравнения несовместны, это означает, что система не имеет точки равновесия и не может являться адекватной моделью реального длительно существующего биоценоза. Потому далее исследование проводим только для четного числа видов.
2. Приведенная система уравнений имеет вид
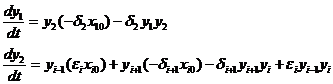 (1-2)
(1-2)
при i=2,3,.., n; δn+1=0
3. Линеаризация системы (1-2) заключается в удалении билинейных членов уравнений (произведений yiyj), а для линеаризованной системы характеристическое уравнение записывается в виде

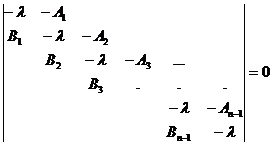
где Ai=δi+1xi0, Bi=εi+1xi+1,0.
Коэффициенты характеристического уравнения вычисляются по формулам:
a0=1; a1=0; a2=![]() ; a3=0;…
; a3=0;…
a2m=![]() a2m+1=0…, где Ci=AiBi.
a2m+1=0…, где Ci=AiBi.
Так как все коэффициенты ai с нечетным индексом равны нулю, все определители Гурвица равны нулю (в каждом из них есть хотя бы один столбец, состоящий только из нулей). Следовательно, система (1-1) находится на границе устойчивости. Это означает, что моделируемая экосистема не может существовать в природе длительное время, поскольку всегда присутствующие в природе внешние воздействия будут выводить ее из состояния равновесия, увеличивая амплитуду колебаний до тех пор, пока для какого-то из видов (звеньев или уровней) величина xi не пересечет нулевой оси, и тогда структура системы будет нарушена.
1.2.2. Влияние связи с косной компонентой типа "запас" на динамику популяций трофической цепи
Рассмотрим теперь уравнения предложенной автором (Булгакова, 1966) модели сообщества, в котором запас пищи для организмов первого трофического уровня ограничен (например, количество биогенов в морской системе или количество гумуса в почве для наземного сообщества). Количество биогенов x0 составляет нижнее звено, которое пополняется за счет потоков трупов и выделений из всех остальных звеньев, и эти потоки пропорциональны биомассе соответствующих звеньев: ![]() .
.
Это звено называем косной (неживой) компонентой.
Блок-схема такой системы приведена на рис. 1.3b, а система уравнений соответствующей модели имеет вид:
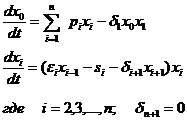 (1-3)
(1-3)
Стационарную точку системы (1-3) находим из уравнений:
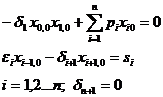 (1-4)
(1-4)
Поскольку уравнение для косной компоненты является нелинейным, процедура определения точки равновесия несколько изменяется по сравнению с вариантом модели Вольтерра: сначала из уравнений n, (n-2), …,(n-2k),… в явном виде находим xn-1,0, xn-3,0,…,xn-2k-1,0,…. Затем последовательно из уравнений (n-1), (n-3), (n-5) и т. д. выражаем
![]()
Подставляя полученные выражения для {xi, o} в уравнение для косной компоненты, получим линейное уравнение относительно xn, o. Таким образом, стационарная точка системы (1-3) существует для любого n и всегда единственна. Чтобы ее координаты были положительными, коэффициенты должны удовлетворять условиям xi,0>0 для всех i.
Приведенная система уравнений модели после линеаризации имеет вид
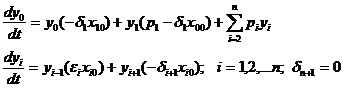 .
.
Подробное исследование, проведенное в работе Т. Булгаковой (1966) с помощью определителей Гурвица, показало, что если существует положительная точка равновесия, первые два определителя Гурвица положительны, остальные приводят к неравенствам над параметрами модели. Выполнение этих условий вместе с условиями {xi,o>0} обеспечивает асимптотическую устойчивость стационарного решения системы.
Рассмотрим, например, случай n=2 (система трех уравнений (1-3)). Координаты стационарной точки равны:
![]() . (1-5)
. (1-5)
Первая координата всегда положительна, а координата косной компоненты x00=(δ2x20+s1)/ε2 положительна, если x20>0. Условие x20>0 может выполняться в двух вариантах:
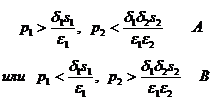 (1-6)
(1-6)
Оказывается, что при выполнении условия A точка равновесия системы уравнений (1-3) положительна и асимптотически устойчива, а при выполнении условия В эта точка неустойчива.
Кроме качественного исследования системы (1-3) на устойчивость, для нескольких вариантов наборов параметров этой модели проведены численные расчеты. Один из примеров численного решения дифференциальных уравнений (1-3) для n=2 показан на рис.1.4. Значения параметров (приведены в подписи к этому рисунку) выбраны так, чтобы они удовлетворяли условию А. Уменьшение коэффициентов ε1 и δ1 означает ослабление связей первого уровня с косной компонентой. Показано, что ослабление этих связей приводит к усилению колебаний, т. е. к понижению устойчивости системы.
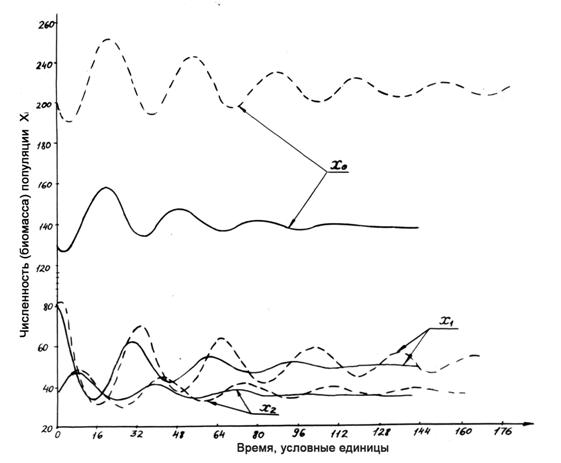
Рисунок 1.4. Динамика компонент модели (1-3) при n=2. Для обоих вариантов δ2=6.78*10-3; ε2=2*10-3; si = σi= 0,1. При ослаблении прямой связи первой компоненты с косной компонентой (при уменьшении коэффициентов δ1 = ε1) колебания усиливаются (пунктирные линии). Сплошной линией показан вариант при δ1 = ε1=2,5*10-3, пунктиром – при δ1 = ε1=1,7*10-3.
Следующий пример (рис.1.5) показывает, что ослабление обратной связи на косную компоненту тоже приводит к усилению колебаний, если это ослабление не слишком велико. Если же, например, порвать обратную связь со второго звена на косную компоненту (p2=0), то второй определитель Гурвица становится равным нулю, поскольку
Δ2=KB0(A0-p1), а A0=p1+p2x20/x10.
Это означает, что при p2=0 система будет находиться на грани устойчивости. На рис. 1.6 показано частное решение этой системы уравнений при начальном значении x0(0)=x00. При р2=0 x0(t)= p1/δ1=const, а система {x1(t),x2(t)} сводится к простой модели Вольтерра (1-1), и динамика этих компонент представляет собой незатухающие колебания возле состояния равновесия.
Еще опаснее нарушение обратной связи с первого уровня (p1=0). При этом x20 <0, т. е. система разрушается.
Рассмотрим некоторое усложнение модели (1-3), делающее ее более реалистичной. В уравнения для каждого i-го трофического уровня (i=1,2,.., n) вводим член, описывающий внутривидовую конкуренцию – зависимость скорости изменения численности i-го уровня от плотности особей этого уровня:
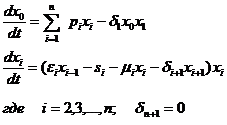 (1-7)
(1-7)
здесь μi- коэффициент внутривидовой конкуренции вида i. При n=2 стационарная точка в общем случае находится решением квадратичного уравнения относительно x20. Просто показать, что при условии x20>0 положительны и другие координаты точки, x10 и x00. Численное решение системы уравнений имеет сначала небольшой выброс, затем без колебаний стремится к положению равновесия (рис. 1.7). Этим подтверждается известный факт, что наличие внутривидовой конкуренции стабилизирует экосистему (Garfinkel, 1967).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 |
