
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
О гидрофобности аминокислот поговорим чуть позже, а для начала полезно увидеть немного цифр, характеризующих гидрофобный эффект при комнатной температуре для неполярных групп, похожих на те, что встречаются в белках.
Таблица 5/1 сразу показывает, что DG (в отличие от DH и DS) растет с размером гидрофобной молекулы.
Таблица 5/1. Характерные термодинамические параметры переноса гидрофобных групп из неполярной жидкости в водный раствор стандартной концентрации (1 моль растворенных молекул на литр) при комнатной температуре (T » 300oK). Данные взяты из C. Tanford, The Hydrophobic Effect (1980).
Вещество | Перенос из ® в | DG | DH | TDS | DCp |
Метан CH4 (ср. с боковой группой Ala: - CH3) | бензол ® вода | +2.6 | -2.8 | -5.4 | +50 |
Бензол C6H6 (ср. с боковой группой Phe: | бензол ® вода | +4.6 | +0.5 | -4.1 | +100 |
Более подробный анализ гидрофобности разных неполярных молекул показал, что их гидрофобная свободная энергия DG растет примерно пропорционально доступной для воды поверхности этих молекул. Смысл и способ построения доступной для воды поверхности демонстрируется рисунком 5-5.
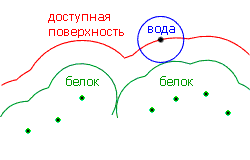
Рис.5-5. Доступная для воды поверхность внедренной в воду молекулы. Точки показывают центры граничащих с водой атомов; жирная линия — их Вандерваальсовы оболочки. Молекула воды представляется как шарик радиусом 1.4 ![]() . "Доступная для воды поверхность" создается центром этого шарика, когда он обкатывает погруженную в воду молекулу, касаясь Вандерваальсовых поверхностей ее внешних атомов. Картинка взята из [3] и адаптирована.
. "Доступная для воды поверхность" создается центром этого шарика, когда он обкатывает погруженную в воду молекулу, касаясь Вандерваальсовых поверхностей ее внешних атомов. Картинка взята из [3] и адаптирована.
Свободная энергия гидрофобного эффекта составляет около +0.02 ккал/моль на каждый квадратный ангстрем доступной поверхности неполярной молекулы, перенесенной из неполярного растворителя в воду. В частности, — для тех молекул, о которых мы уже говорили: для метана — DG » 2.6 ккал/моль при доступной поверхности примерно в 150 ![]() 2; для бензола — DG » 4.6 ккал/моль при доступной поверхности примерно в 200
2; для бензола — DG » 4.6 ккал/моль при доступной поверхности примерно в 200 ![]() 2; для циклогексана — DG » 6 ккал/моль при доступной для воды поверхности примерно в 300
2; для циклогексана — DG » 6 ккал/моль при доступной для воды поверхности примерно в 300 ![]() 2.
2.
Та же закономерность наблюдается и для гидрофобных аминокислотных остатков (Рис.5-6).
У боковых групп с полярными атомами гидрофобный эффект (при той же общей доступной для воды поверхности) меньше. Однако, если взять только ту доступную для воды поверхность, которая создается неполярными атомами (т. е. вычесть из общей доступной для воды поверхности приблизительно по 50 ![]() 2 на каждый полярный атом), — мы получим примерно ту же зависимость гидрофобности от размера доступной для воды неполярной поверхности боковой группы.
2 на каждый полярный атом), — мы получим примерно ту же зависимость гидрофобности от размера доступной для воды неполярной поверхности боковой группы.
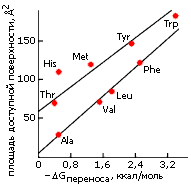
Рис.5-6. Размер доступной для воды поверхности боковых цепей аминокислотных остатков и гидрофобность этих боковых цепей. [Доступная поверхность боковой цепи остатка Х равна доступной поверхности аминокислоты Х за вычетом доступной поверхности остатка Gly, не имеющего боковой цепи; гидрофобность боковой цепи остатка Х равна экспериментально измеренной гидрофобности остатка Х минус гидрофобность остатка Gly]. Остатки Ala, Val, Leu, Phe имеют чисто углеводородные боковые цепи. У Thr и Tyr в боковой цепи есть еще по одному О-атому, у Меt — S-атом, у Trp — N-атом, у His — два N-атома; поэтому у всех них размер доступной неполярной поверхности меньше полного размера доступной поверхности. Картинка взята из [3].
Гидрофобный эффект играет очень важную роль в поддержании стабильности белковой структуры. Именно он сворачивает белковую цепь в компактную плотную глобулу. Рисунок 5-7 показывает, что свободная энергия перехода гидрофобной группы из воды в гидрофобную же жидкость велика — она составляет несколько килокалорий на моль, в то время как свободная энергия затвердевания этой же неполярной жидкости близка к нулю при физиологических температурах. [Точнее: Для сравнительно низкомолекулярных углеводородов, — таких, как циклогексан, показанный на Рис.5-7, — свободная энергия отвердевания даже положительна, и это не дает им отвердеть при комнатной температуре. Отвердеванию противится энтропия, в частности — энтропия вращений и перемещений молекул по жидкости, — в жидкости каждая молекула плавает более или менее свободно, а в твердом теле она зажата кристаллической решеткой. Энтропия перемещений молекулы, грубо говоря, не зависит от ее размера — в отличие от энтальпии, которая растет с ростом числа контактов одной молекулы с другими, т. е. с ростом ее поверхности. В белковой цепи роль энтропии перемещений меньше, так как аминокислотные остатки связаны цепью, т. е. не могут двигаться независимо друг от друга, и это способствует отвердеванию белка.]
Если даже пренебречь всей вообще энтропией кристаллизации циклогексана в свободной энергии его отвердевания, DGжидкий_циклогексан-Solid, т. е. положить DGжидкий_циклогексан-Solid » DH жидкий_циклогексан-Solid = —2 ккал/моль, — то и в этом случае видно, что для циклогексана термодинамический эффект отвердевания много слабее гидрофобного эффекта, приводящего к конденсации растворенных в воде молекул в жидкую каплю, так как DGвода-жидкий_циклогексан = —6 ккал/моль.
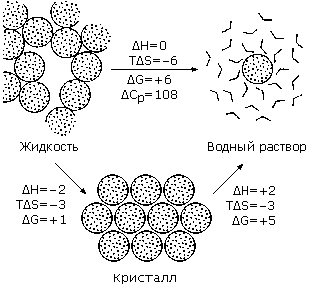
Рис.5-7. Термодинамика переноса типичной неполярной молекулы, циклогексана, из жидкой фазы в твердую и в водный раствор. Цифры соответствуют процессам при 25оС. Величины DH, ТDS и DG даны в ккал/моль. Термодинамические величины для жидкости и твердого тела относятся к стандартному давлению 1 атм., а раствора — к его стандартной концентрации 1 моль/литр. Картинка взята из [6] и адаптирована.
Таким образом, гидрофобный эффект берет на себя, грубо говоря, три четверти работы по созданию белковой глобулы. Но он сам по себе не может создать нативный твердый белок. Он создает лишь расплавленную белковую глобулу, о которой речь впереди. Отвердевает белок, как и все органические жидкости, под действием Вандерваальсовых сил, а также водородных и ионных связей — взаимодействий более специфичных и более чувствительных к деталям атомной структуры, чем простая водобоязнь. Но это — окончательная огранка, а большая часть работы, вся черновая работа падает на гидрофобный эффект.
Лекция 6
Сегодня мы обсудим электростатические взаимодействия, точнее — те особенности, которые им придает наличие белковой глобулы и воды вокруг нее.
Казалось бы, что здесь обсуждать: вы и так помните, заряд q1 на расстоянии rот себя создает, в среде с диэлектрической проницаемостью e, электрическое поле с потенциалом
j = q1/er , | (6.1) |
и взаимодействует с находящимся на этом расстоянии зарядом q2 с энергией
U = jq2 = q1q2 /er . | (6.2) |
Помните вы, наверно, и то, что в вакууме (или в воздухе) e = 1, в воде — e близко к 80, а в среде типа пластмассы (и, между прочим, сухого белка) e лежит где-то между 2 и 4.
Это все верно, — но только "почти". Во-первых, U является, строго говоря, не энергией, а свободной энергией (напомню, что именно свободная энергия стремится к минимуму при приближении к равновесию, т. е. именно она должна отвечать, в теплой среде, за притяжение разноименных зарядов и отталкивание одноименных). Но это — постольку, поскольку нас интересует влияние взаимодействия зарядов на стабильность белка — мелочь, пуристика. Важнее то, что уравнение (6.2) справедливо только тогда, когда "среда" однородна. А когда нас интересует взаимодействие зарядов в белках, мы имеем дело с крайне неоднородной средой. У самог![]() белка, как у любой пластмассы, e невелико, где-то около 2 или 4 (напомню: в воздухе e=1). А у воды e=80. И заряженные группы белка обычно расположены на его поверхности, у самой воды (позже мы поймем, почему). Какое же e мы должны брать для оценок электростатических взаимодействий в белке? Если мы выберем e»80, два единичных (протонных) заряда на расстоянии 3
белка, как у любой пластмассы, e невелико, где-то около 2 или 4 (напомню: в воздухе e=1). А у воды e=80. И заряженные группы белка обычно расположены на его поверхности, у самой воды (позже мы поймем, почему). Какое же e мы должны брать для оценок электростатических взаимодействий в белке? Если мы выберем e»80, два единичных (протонных) заряда на расстоянии 3 ![]() будут взаимодействовать с энергией »1.5 ккал/моль; если же мы возьмем e»3, то они будут взаимодействовать с энергией »40 ккал/моль. Разница уж очень велика: дополнительные +40 ккал/моль разрушат любую белковую структуру...
будут взаимодействовать с энергией »1.5 ккал/моль; если же мы возьмем e»3, то они будут взаимодействовать с энергией »40 ккал/моль. Разница уж очень велика: дополнительные +40 ккал/моль разрушат любую белковую структуру...
Второй вопрос связан вот с чем. Формулы (6.1 и 6.2) справедливы, когда r, расстояние между зарядами, много больше размера молекул среды. Но в белках заряды часто находятся в прямом контакте, на расстоянии 3-4 ![]() друг от друга, когда даже молекула воды, не говоря уже о боковой группе, между ними не влезет. Как же оценивать электростатические взаимодействия в этом случае? Брать e=1, как в вакууме, что ли?
друг от друга, когда даже молекула воды, не говоря уже о боковой группе, между ними не влезет. Как же оценивать электростатические взаимодействия в этом случае? Брать e=1, как в вакууме, что ли?

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
