
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
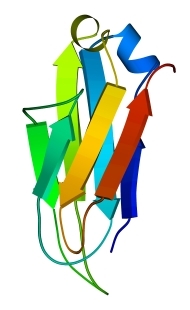
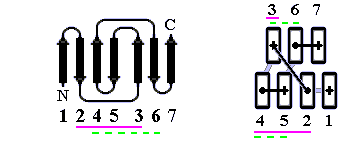
Рис.13-9. Продольная упаковка b-листов в константном домене легкой цепи иммуноглобулина ![]() . Слева представлена подробная картина белка; радужная расцветка (от синего до красного) трассирует ход цепи, от N - к С-концу. На топологической схеме (в центре рисунка) подчеркнуты "греческие ключи". На рисунке справа дан вид на белок "снизу" (с торцов структурных сегментов). Прямоугольники - торцы b-тяжей. Крестик соответствует N-концу сегмента (т. е. он "идет от нас"), точка — его С-концу (т. е. он "идет к нам"). Ход петель, соединяющих структурные сегменты, показан черной линией, если петля обращена к нам, и светлой, если она находится на противоположной стороне укладки. Обратите внимание, что такая схема позволяет представить колинеарную упаковку этих сегментов (b-тяжей) наиболее просто. Кроме того, она дает возмохность увидеть пространственное строение "греческих ключей" и заметить, что два имеющихся "ключа" в пространстве организованы по-разному.
. Слева представлена подробная картина белка; радужная расцветка (от синего до красного) трассирует ход цепи, от N - к С-концу. На топологической схеме (в центре рисунка) подчеркнуты "греческие ключи". На рисунке справа дан вид на белок "снизу" (с торцов структурных сегментов). Прямоугольники - торцы b-тяжей. Крестик соответствует N-концу сегмента (т. е. он "идет от нас"), точка — его С-концу (т. е. он "идет к нам"). Ход петель, соединяющих структурные сегменты, показан черной линией, если петля обращена к нам, и светлой, если она находится на противоположной стороне укладки. Обратите внимание, что такая схема позволяет представить колинеарную упаковку этих сегментов (b-тяжей) наиболее просто. Кроме того, она дает возмохность увидеть пространственное строение "греческих ключей" и заметить, что два имеющихся "ключа" в пространстве организованы по-разному.
Подобным же образом уложены цепи белков примерно пятидесяти (!) других суперсемейств, — белков, не похожих на иммуноглобулин по аминокислотной последовательности (хотя часть из них и отвечает, как иммуноглобулин, за специфическое связывание каких-то объектов — например, при межклеточном узнавании). Помимо удовольствия показать вам столь популярный мотив укладки цепи, я преследую еще одну цель: показать, что мотив строения белка с более или менее колинеарной упаковкой структурных сегментов проще всего отражается схемой, представляющей вид с торца на такой белок.
Перейдем к ортогональной упаковке b-листов. Рисунок 13-10 показывает, как один и тот же b-цилиндр (при различной топологии, при различном ходе цепи через ортогональную укладку b-листов) служит основой и сериновой протеазы типа трипсина (а), и протеазы кислой, типа пепсина (б).
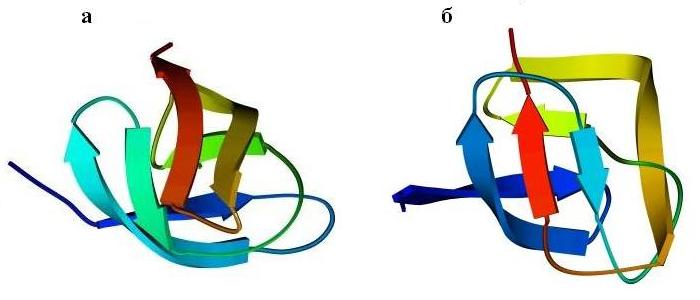
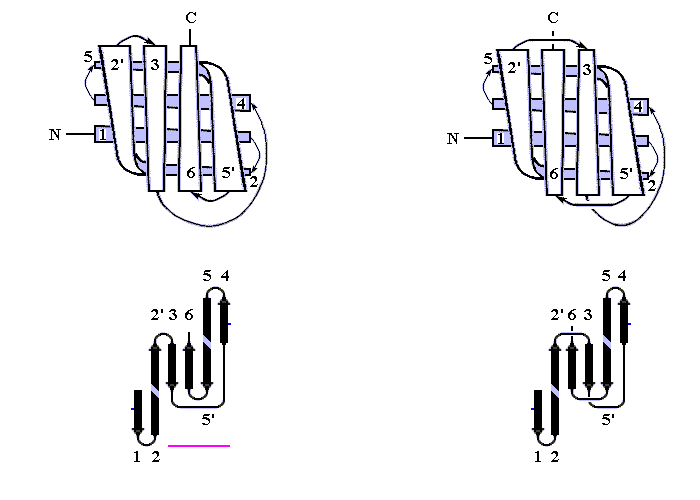
Рис.13-10. Мотив укладки цепи в сериновой протеазе (а) и в кислой протеазе (б). В последнем белке петли укорочены и изображены довольно схематично. Показана схема ортогональной упаковки b-листов в этих белках, а также топологические схемы b-листов. Эти листы согнуты (при изгибе края листов уходят от читателя и смыкаются; возникающая при этом водородная связь краев b-листа изображена черточками); места перегибов b-тяжей соответствуют их более светлым частям. "Греческий ключ" в трипсине подчеркнут.
При этом b-домен белка — катаболитического активатора имеет точно такой же мотив укладки цепи (т. е. ту же топологию хода цепи через такого же вида b-структурный штабель), что и совершенно не связанный с ним (ни генетически, ни функционально) белок оболочки сателлитного вируса некроза табака.
Примеры такого сходства структур при отсутствии какой-либо другой видимой связи между белками очень многочисленны.
Выше мы рассмотрели "базовые" конструкции b-белков. Есть и более сложные — например, "многозаходный пропеллер".
В "пропеллере" нейраминидазы (Рис.13-11) — шесть наклонно уложенных b-листов образуют розетку (в других белках бывает до восьми b-листов). Если рассматривать эти листы попарно — они образуют b-сэндвичи, так что "пропеллер" часто описывают как суперцилиндр, сложенный из b-сэндвичей.
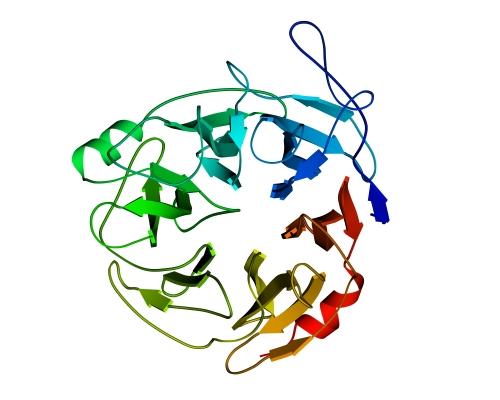
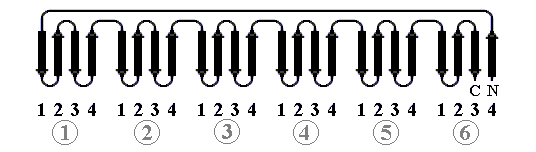
Рис.13-11. b-Структура в форме "шестилопастного пропеллера" в нейраминидазе. Внизу — схема топологии этого белка, состоящего из шести антипараллельных b-листов.
Во "вмятине" на оси этого суперцилиндра — вы видите, что она не прикрыта петлями — находится активный центр. В ретинол-связывающем белке мы уже встречались с таким расположением активного центра — во вмятине в центре цилиндра, и мы встретимся с ним еще.
Интересна также — прежде всего своей регулярностью — конструкция типа "b-призмы" или (другое название) "b-спирали" (Рис.13-12). Три грани такой призмы образованы тремя b-листами — причем параллельными! — а цепь проходит сквозь них по спирали, непрерывно переходя из одного листа в другой. При этом она как бы навивается на ось призмы, образуя либо обычную при соединении параллельных b-тяжей правую спираль, либо (в других призмах) спираль левую, исключительно редкую при соединении b-тяжей.
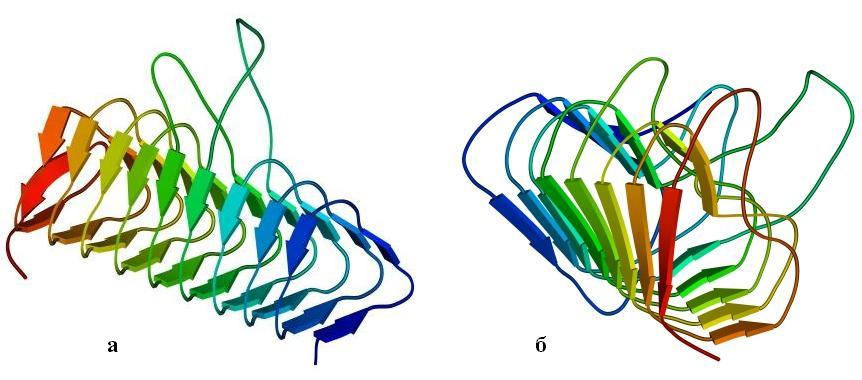
Рис.13-12. b-Призма в ацилтрансферазе (а) и в пиктатлиазе С (б). Обратите внимание на разную закрученность цепи вокруг длинной оси призмы: необычную, левую на рисунке (а) и обычную, правую — на рисунке (б), а также на то, что при левой закрученности цепи отсутствует обычная (правопропеллерная, если смотреть вдоль b-тяжей, см. Рис.13-12б и Рис.13-5 — 13-11) скрученность b-листа.
Теперь уместно поговорить о топологии b-белков. Прежде всего обращает на себя внимание то, что b-белки сложены преимущественно из антипараллельной b-структуры. Большинство рассмотренных нами до сих пор b-белков состояло из чистой антипараллельной b-структуры. Порой к ним примешивается небольшое количество параллельной (см. Рис.13-10б). Очень редко, но бывают белки, сложенные из чисто параллельной b-структуры (см. Рис.13-12).
То, что "примесь" параллельной к антипараллельной b-структуре довольно мала — в общем, не удивительно, так как они имеют несколько разные конформации, так что их стык должен быть энергетически невыгодным. В какой мере энергетическая невыгодность структуры сочетается с относительной редкостью ее встречаемости — об этом мы поговорим на одной из ближайших лекций; однако в целом ясно, что стабильная система — а белок стабилен — должна состоять в основном из стабильных же элементов и избегать внутренне нестабильных.
Преимущественно же антипараллельный характер b-листов в b-белках тесно связан с тем, что их архитектура обычно основана на b-шпильках (Рис.13-13). Такие шпильки часто бывают согнуты пополам, а иногда — согнуты дважды или даже трижды (как на Рис.13-8б, в).

Рис.13-13. Антипараллельные b-шпильки.
Петли, соединяющие b-участки, обычно имеют вход и выход на одном и том же краю укладки (т. е. они не пересекают "штабель", а прикрывают его торец). Это видно почти на всех рисунках. При этом петли, даже если они длинны, чаще всего соединяют близкие между собой в пространстве концы b-участков. Поэтому соседние по цепи b участки, как правило, не параллельны — и часто образуют анитипараллельные b-шпильки.
Отметим также, что "наложение" петли на петлю (или, как говорят, "пересечение петель") встречается редко (такое исключение из правил показано на Рис.13-10б, снизу), — видимо, потому что при этом петли должны дополнительно изгибаться (чтобы избежать столкновения или дегидратации), что опять-таки энергетически невыгодно. Избегание наложения петель — общее структурное правило, наблюдающееся в белках.
В результате, из многих изображенных на Рис.13-14 возможных конфигураций b-листа, сложенного из единого куска цепи, — по-настоящему часто встречаются лишь три мотива: "меандр" и два "греческих ключа" (они подчеркнуты на рисунке) — именно эти мотивы свободны от всех отмеченных выше недостатков.
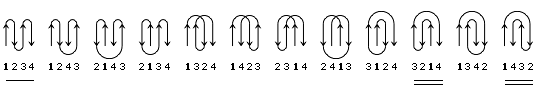
Рис.13-14. Возможные топологии листов из четырех b-участков. Показаны только листы, где каждые два подряд (в цепи) идущих b-участка направлены в противоположные стороны. Из этих топологий часто встречаются только "меандр" (подчеркнут одной чертой) и два (подчеркнуты двойной чертой) "греческих ключа" (они различаются только направлением поворота цепи от шпильки из тяжей 1,4 к шпильке из тяжей 3,4). Пример белка с меандром — ретинол-связывающий белок (см. Рис.13-7); примеры белков с "греческим ключом" — g-кристаллин и другие белки, изображенные на Рис.13-8), или трипсин (Рис.13-10).
"Меандр" — это, кстати, имя очень извилистой реки в Греции — характерен тем, что соседние по цепи b-тяжи являются также ближайшими соседями в пространстве (Рис.13-8); обычно (но не всегда) они связаны водородными связями. "Греческий ключ" (такой орнамент можно увидеть на старых вазах и садовых решетках) характерен тем, что четыре соседних по цепи b-тяжа антипараллельны, причем первый и четвертый связаны водородными связями. При этом на самом деле второй и/или третий b-тяжи "ключа" часто лежат не в том же (как может показаться из Рис.13-14), а в другом b-листе. При этом в пространстве образуются различные структуры — так называемые "abcd" структуры Ефимова — с одной и той же топологией греческого ключа (Рис.13-15). Найдите их на рисунках 13-8, 13-10.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
