
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
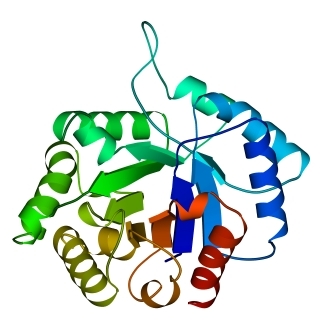
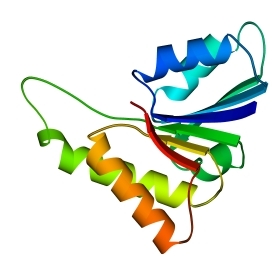
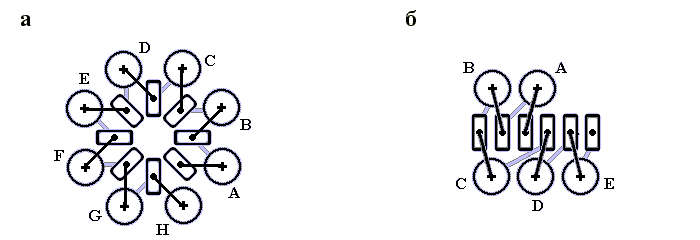
Рис.14-11. Типичные мотивы строения a/b белков и их упрощенные модели (вид на модели — с торца b-слоя): "a/b цилиндр" в триозофосфатизомеразе (а); "укладка Россманна" в NAD-связывающем домене малатдегидрогеназы (б). На детальной картинке в первом белке видна обращенная к нам впадина, образованная расходящимися в форме розетки петлями; она идет к центру b-цилиндра. Во втором — слева вверху видна щель между расходящимися вниз и вверх петлями.
В a/b цилиндре обычно насчитывается восемь a - и восемь b-участков, причем почти все a/b цилиндры имеют одинаковую топологию: все соседние и расположенные через один по цепи b - и a-участки контактируют друг с другом. По-видимому, такое строение обеспечивает особую стабильность белковой глобулы, так как многочисленные белковые глобулы с такой архитектурой (наблюдаемой в 10% белков) все очень похожи друг на друга по форме, — часто и без каких-либо следов общего происхождения в аминокислотной последовательности, и без какой-либо общности функций.
Без общности функций - да; без общности в строении активного центра - да; но не без общности места активного центра в архитектуре глобулы, — каждая архитектура содержит места (обычно — впадины), как бы специально созданные этой архитектурой для активного центра — что бы он ни делал.
Я хочу обратить ваше внимание на "воронку" на оси a/b цилиндра (Рис.14-12а), — вы видите, что эта вмятина, заложенная в общей архитектуре белка, не прикрыта петлями. Здесь находится активный центр. Точнее: из двух таких "воронок", расположенных на противоположных торцах a/b цилиндра, под активный центр используется только одна, — та, куда смотрят С-концы b-участков и N-концы a-спиралей. Считается, что именно эти концы (возможно, из-за множества открытых NH-групп N-концов спиралей), вместе с соединяющими их короткими петлями, особенно пригодны для связывания разнообразных субстратов. Впрочем, этот вопрос еще недостаточно ясен.
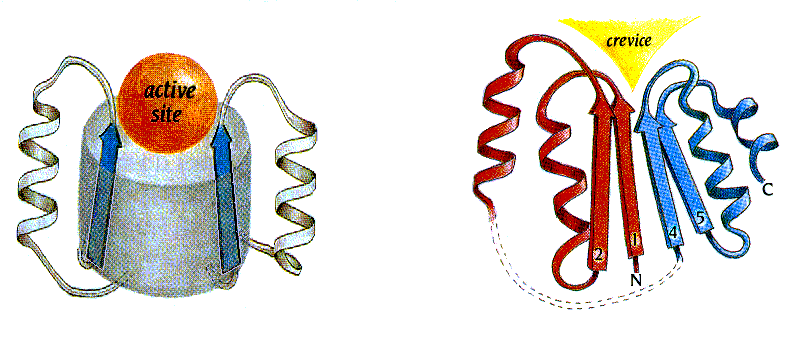
Рис.14-12. Типичное положение активного центра (active site) в a/b белках: в "воронке" на оси a/b цилиндра (а), и в щели (crevice), образованной расходящимися петлями в "укладке Россманна" (б). Картинки, с небольшими изменениями, взяты из [5].
Аналогичное размещение активного центра — в щели, во вмятине, причем во вмятине, куда тоже смотрят С-концы b-участков и N-концы a-спиралей — наблюдается и в "укладках Россманна". Только здесь вмятина образуется не при расхождении петель от центра цилиндра, а при расхождении петель, одна часть которых идет от b-листа к спиралям лежащим под листом, а другая — над этим листом (Рис.14-12б).
Теперь перейдем к a+b белкам. В их основе лежит антипараллельная (а не параллельная, как в a/b белках) b-структура.
В a+b белках выделяются два класса. Белки одного класса (их порой называют ab-складками) напоминают a/b белки тем, что в них слой a-спиралей лежит на b-листе. Они напоминают a/b белки также регулярным (но с иным, чем в a/b белках) чередованием a и b участков в цепи и в пространстве. Белки другого класса ("собственно" a+b белки) не имеют такого чередования, в их цепях a-структура "отмешана" от b-структуры.
Характерное чередование a - и b-участков в цепи ab-складки имеет вид...a-b-b-a-b... или......a-b-b-b-b-a-b-b... (Рис.14-13). Здесь отдельные a-спирали лежат между b-шпильками или b-листами из четного числа b-тяжей. Соседние по цепи b-участки в ab-складках образуют антипараллельные b-шпильки; а из-за четного (а не нечетного, как в a/b белках) числа b-тяжей между a-спиралями, и из-за общей колинеарности этих тяжей a-спиралям, — a-спирали также образуют антипараллельные шпильки. Интересно, что такая (или, точнее, сходная — abbabb) "складчатая" конструкция белка была сначала теоретически предсказана (точнее — предложена в качестве проекта белково-инженерного дизайна), а затем уже такие архитектуры были массово обнаружены в природе, — причем их оказалось особенно много среди РНК-связывающих белков.
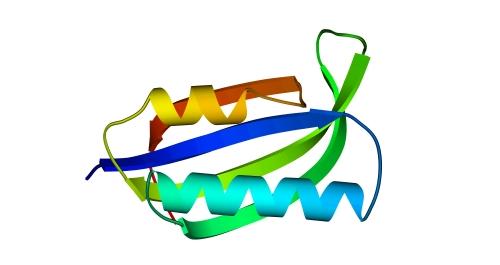
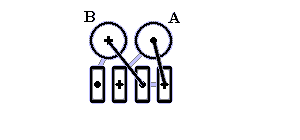
Рис.14-13. Один из типичных мотивов строения a+b белка: "ab складка" (ab-plait) в рибосомальном белке S6. ab складка отличается более регулярным, чем в "собственно" a+b белках, чередованием вторичных структур в цепи (в данном случае: babbab). Радужная кодировка (синий - голубой - зеленый - желтый - оранжевый - красный) позволяет проследить ход цепи, от N - к С-концу. Справа помещена схема строения этого белка (вид с торца структурных участков, приблизительно колинеарных). Спирали занумерованы буквами. "+" на торце a - или b-участка означает, что он идет от нас (т. е. что к нам обращен его N-конец), точка — что к нам.
В "собственно" a+b доменах (Рис.14-14) a - и b-участки расположены в цепи нерегулярно, и скорее — как бы блоками. Такие белки обычно выглядят как b-лист (часто — загнутый сам на себя), прикрытый отдельными a-спиралями или a-спиральным субдоменом; b-структура в них в основном антипараллельна, как в "чистых" b белках.
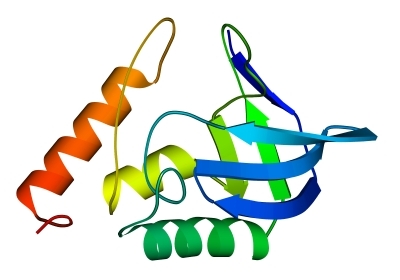
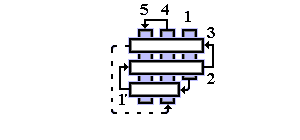
Рис.14-14. Характерный мотив строения a+b белков: нуклеаза стафиллококка. "Собственно" a+b белки отличаются менее регулярным, чем в a/b белках или ab-складках, чередованием вторичных структур в цепи (в данном случае: bbbabbaa). Мотив укладки цепи, наблюдаемый в b-домене нуклеазы, называется "ОБ-укладка" ("OB-fold", то есть "Oligonucleotide-Binding fold", или "укладка [имени] Олега Борисовича [Птицына]"). Справа помещена схема строения этого, часто встречающегося в разных белках, ОБ-домена (вид сверху на ортогональную упаковку b-участков). b-тяжи занумерованы цифрами.
Очень характерной чертой a/b и a+b белков (а равно и b-белков) является правовинтовой (т. е. "к нам — против часовой стрелки") ход перемычек (Рис.14-15) между параллельными b-участками одного и того же b-листа, следующими друг за другом в цепи (но не обязательно при том соседними в этом листе) (см. Рис.14-11 и далее). При этом перемычка между такими параллельными b-участками в a/b - и a+b-белках, как правило, включает в себя a-спираль. В b (и порой в a+b) белках в такой правовинтовой перемычке между параллельными b-участками одного листа лежит — вы должны помнить из прошлой лекции ("abcd" структуры и т. д.) b-участок другого листа, а иногда и отдельный b-лист. Иногда (довольно редко) в перемычке между параллельными b-участками нет ни a-, ни b-структуры, — но и тогда эта перемычка обычно право-, а не левовинтовая.
На следующей лекции мы увидим, что такой ход перемычки позволяет большее разнообразие кодирующих, т. е. стабилизирующих такую структуру последовательностей.
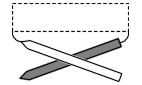
Рис.14-15. Типичный, правовинтовой ход перемычек между параллельными b-тяжами одного листа. В перемычке обычно находится еще один участок вторичной структуры.
Завершая обзор структур глобулярных белков, я хочу еще раз подчеркнуть, что одни и те же — или очень похожие — архитектуры часто встречаются в белках, совсем не сходных функционально или филогенетически. Это открытие заложило основы физической, или рациональной (как это чаще называется в литературе) классификации белков.
Наиболее полная и популярная на сей день классификация представлена в компьютерном классификаторе SCOP (Structural Classification of Protein), сделанном после его переезда из Пущино в Кембридж. Она начинается с класса белка (a, b и т. д.), классы подразделяются по мотивам укладки цепи, и уже мотивы подразделяются на суперсемейства, где просматривается хоть какая-то гомология последовательностей, те — на семейства с явно проявляющейся гомологией, и т. д. Но, пожалуй, еще более последовательно физическая классификация белковых структур проводится классификатором САТН (Class - Architecture - Topology - Homology), сделанном в группе Дж. Торнтон в Лондоне.
Лекция 15
Теперь поговорим об общих закономерностях, наблюдаемых в структуре белков. Из прошлых лекций вы должны были вынести впечатление, что большинство белковых цепей вписывается в узкий набор стандартных структур.
На самом деле здесь действует правило "80%:20%". В его исходном виде оно гласит: "80% всего пива выпивается 20% населения". В применении к белкам — "80% всех белков вписывается в 20% наблюдаемых архитектур белковых глобул". И я позволил себе сосредоточиться именно на типичных структурах.
Вопрос — почему же большинство белков вписывается в узкий набор стандартных структур? И почему не все (как цепи ДНК)? На каком структурном уровне проявляется это сходство? И что стоит за этими общими структурами: память об общем происхождении? функциональная целесообразность? или необходимость удовлетворять общим принципам сворачивания стабильных белковых структур?

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
