
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Более точно оценить величину e на самых малых близких расстояниях внутри молекулы можно, исходя из величин первой и второй констант диссоциации двухосновных кислот и оснований в воде. Например, диссоциация щавелевой кислоты происходит так:
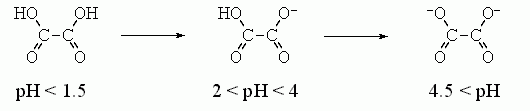
Вторая диссоциация сдвинута на »2.5 единицы рН относительно первой, т. е. происходит при в 102.5 = е2.3x2.5 раз меньшей концентрации Н+. Это показывает, что свободная энергия взаимодействия первого заряда со вторым составляет 2.5x2.3RT » 3.5 ккал/моль. И это — при расстоянии в »3 ![]() между зарядами. Такая величина энергии взаимодействия соответствует e»30?40 на расстоянии в 3
между зарядами. Такая величина энергии взаимодействия соответствует e»30?40 на расстоянии в 3 ![]() . К сходному результату — e» 30?40 на расстоянии в » 2-2.5
. К сходному результату — e» 30?40 на расстоянии в » 2-2.5 ![]() — приводит рассмотрение диссоциации угольной кислоты, Н2СО3® НСО3-® СО3--.
— приводит рассмотрение диссоциации угольной кислоты, Н2СО3® НСО3-® СО3--.
Значит, даже солевая связь противоположно заряженных боковых групп на поверхности белка должна "стоить" всего пару-тройку ккал/моль. Внутри белка она должна "стоить" больше, но погружение заряженных групп внутрь глобулы обойдется еще дороже — так что не надо удивляться, что такие связи в природных белках наблюдаются нечасто.
Итак, мы пришли к выводу, что корпускулярность не меняет радикально "макроскопическую" (выведенную для больших расстояний между зарядами) диэлектрическую проницаемость воды даже на расстоянии в 2 — 3 ![]() , когда между взаимодействующими зарядами не может быть никаких других молекул. Причина этого — в том, что заряды достаточно сильно экранируются даже подошедшими "с другой стороны и с боков" (Рис.6-13) молекулами среды, которые поляризуются (в случае воды — просто поворачиваются) так, что "+"'ы этих молекул смещается к заряду
, когда между взаимодействующими зарядами не может быть никаких других молекул. Причина этого — в том, что заряды достаточно сильно экранируются даже подошедшими "с другой стороны и с боков" (Рис.6-13) молекулами среды, которые поляризуются (в случае воды — просто поворачиваются) так, что "+"'ы этих молекул смещается к заряду ![]() , а "-"'ы — к заряду
, а "-"'ы — к заряду ![]() .
.
| Рис.6-13. |
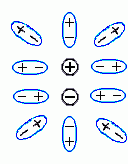
Здесь мы снова (ср. Рис.6-10, 6-11) видим, что электростатическое взаимодействие между зарядами (![]() и
и ![]() ) как бы проходит в основном через среду с более высокой диэлектрической проницаемостью и почти игнорирует среду, которая слабо поляризуется.
) как бы проходит в основном через среду с более высокой диэлектрической проницаемостью и почти игнорирует среду, которая слабо поляризуется.
Все, о чем мы говорили до сих пор, относилось к "мелкомолекулярным" системам. Сохраняются ли все эти выводы для белков?
Опыты, проведенные в лаборатории Фершта — зачинателя белковой инженерии — показывают, что приведенные выше оценки в полной мере относятся и к белкам.
Опыты основывались на следующем. Есть белки, ферменты, активность которых особенно велика при определенном значении рН (у них, как говорится, есть рН-оптимум).
Вводя в белок, путем мутации его гена, заряженный остаток, можно сдвинуть этот рН-оптимум (Рис.6-14) — и, по его сдвигу, оценить электрическое поле, созданное мутировавшим остатком в активном центре.
| Рис.6-14. |
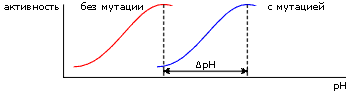
Причина рН-оптимума — в том, что какая-то группа в активном центре должна, для успешной работы фермента, находиться в определенной зарядовой форме, а заряженность этой группы зависит от концентрации водородных ионов в среде.
Пусть активный центр (АЦ) принимает ион Н+: АЦ + Н+ = АЦН+. Тогда, по закону действующих масс, соотношение концентраций двух (с Н+ и без Н+) форм активного центра составляет [АЦН+]/[АЦ] = exp(-DFАЦН+/RT) x [H+] = exp(-DFАЦН+/RT) x 10-рН = exp{-(DFАЦН+/RT + 2.3 x рН)}, где DFАЦН+ — свободная энергия связывания Н+ с активным центром, а значок [ ] означает концентрацию.
Если мутация вносит в белок заряд, создающий в его активном центре потенциал j, то DF АЦН+ меняется: DF АЦН+|с_мутацией = DFАЦН+ |без_мутации + je, где e — заряд Н+. Так как в рН-оптимуме величина [АЦН+]/[АЦ] (и, следовательно, величина DFАЦН+/RT + 2.3 x рН) должна быть одной и той же и при, и без мутации, — то DFАЦН+|без_мутации /RT + 2.3 x рН|опт._без_мутации = DFАЦН+|с_мутацией /RT + 2.3 x рН|опт._с_мутацией. То есть je = DFАЦН+|с_мутацией — DF АЦН+|без_мутации = 2.3RT(рН|опт._без_мутации — рН|опт._с_мутацией) = 2.3RTx(-DрН).
Так, зная сдвиг pH-оптимума, можно оценить потенциал, созданный в активном центре мутированным остатком белка. Теперь, зная трехмерную структуру этого белка и, следовательно, расстояние r от мутированного остатка до активного центра, — можно оценить эффективную диэлектрическую проницаемость eэфф (входящую в формулу j = q/eэффr) для взаимодействия внесенного мутацией заряда q с районом активного центра.
Мутации в опытах Фершта вводились на поверхность белка, чтобы не разрушить его структуру (мы уже знаем, что энергия глубоко погруженного в белок заряда очень велика, так что он может буквально взорвать белок).
Результат: эффективная диэлектрическая проницаемость eэфф колебалась в этих опытах от ~40 до ~120, причем первое характерно для близких к активному центру мутаций, а второе — для удаленных от него. Последнее — то, что eэфф может достигать 120 — вызвало изрядное удивление, так как многие, не знакомые как следует с электростатикой, считали, что eэфф должны лежать где-то между 3 (как внутри белка) и 80 (как в воде). Однако нас эти величины удивлять не должны — они хорошо согласуются с тем, что можно ожидать из Рис.6-10.
Небольшое отступление — о белковой инженерии. Ее главная прелесть заключается в том, что, меняя такой-то кодон в гене белка, мы можем ввести мутацию в точно определенное место белковой глобулы, так как и ген этого белка, и его аминокислотная последовательность, и его трехмерная структура известны. Кроме того, влияние мутации на структуру также можно контролировать — рентгеноструктурным анализом или ЯМР. Таким образом, вся работа ведется с открытыми глазами.
В опытах, о которых шла речь, белок служит микроскопическим (вернее, наноскопическим) электрометром. И белковая инженерия дает возможность использовать такие приборы — и при этом прыгать от физической теории к генным манипуляциям и обратно, что чрезвычайно занимательно.
Теперь я хочу сделать несколько добавлений касательно электростатических взаимодействий.
Первое. До сих пор я говорил только о взаимодействии отдельных зарядов. Однако к электростатике относятся и взаимодействия диполей (например, диполей Н(+)-О(-) и Н(+)-N(-), вовлеченных в образование водородных связей), а также квадруполей — последние присутствуют, например, в ароматических кольцах (Рис.6-15).
Рис.6-15. Электрический квадруполь ароматического кольца: |
|
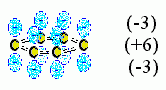
Я остановился на взаимодействии зарядов потому только, что они наиболее сильны, — даже при прямом контакте они в несколько раз сильнее, чем взаимодействия диполей (и к тому же медленнее спадают с расстоянием), а взаимодействия диполей, — сильнее, чем взаимодействия квадруполей.
Второе. При наличии свободных зарядов (например, соли) в воде, электростатические взаимодействия ослабевают с расстоянием r не по закону "энергия пропорциональна (1/r)", а гораздо быстрее — как (1/r) x exp(-r/D). Здесь D — радиус Дебая-Хюккеля — соответствует характерному размеру противоионного облака вокруг заряда. Величина D не зависит от самого заряда, но зависит от концентрации ионов в среде, от ее диэлектрической проницаемости и температуры. В воде, при комнатной температуре,
D » 3/I1/2 | (6.5) |
где
I = 1/2 åi ci zi2 — | (6.6) |
ионная сила раствора в моль/литр. В формуле (6.6) сумма берется по всем сортам ионов в растворе, причем zi — заряд (в единицах протонного заряда), а ci — концентрация (в моль/литр) иона i. Обычным физиологическим условиям соответствует I » 0.1-0.15; при этом D » 8![]() . Однако некоторые микроорганизмы живут при I ~ 1 моль/л и выше; при этом сохраняются, и то в очень ослабленном виде, только те электростатические взаимодействия, что соответствуют "солевым мостикам", т. е. прямому контакту зарядов.
. Однако некоторые микроорганизмы живут при I ~ 1 моль/л и выше; при этом сохраняются, и то в очень ослабленном виде, только те электростатические взаимодействия, что соответствуют "солевым мостикам", т. е. прямому контакту зарядов.
В целом, при наличии в растворе ионной атмосферы, энергия взаимодействия двух зарядов имеет вид

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
