
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рис.19-16. (а) Структура исходного "цинкового пальца" (второго модуля белка Zif268); ион Zn показан шариком. (б) Структура искусственного белка FSD-1. (в) Спектр КД для FSD-1 при 1оС. (г) Изменение спектра КД искусственного FSD-1 с температурой. Картинки взяты из B. I.Dahiyat & S. L.Mayo, Science (1997) 278:82-86.
Кроме белков, основанных на природных архитектурах, конструируются искусственные белки, чьи архитектуры не имеют природных аналогов.
В основу искусственного белка альбебетина (Рис.19-17а) положена структура, не обнаруженная еще в природе. Она состоит из двух повторов типа a-b-b (поэтому белок и назван альбебетином), и она не противоречит общим принципам формирования структуры глобулярных белков. Вместе с , я занимался дизайном альбебетина, а получение и исследование этого белка и его вариантов велось здесь, в Институте белка, и в группе .
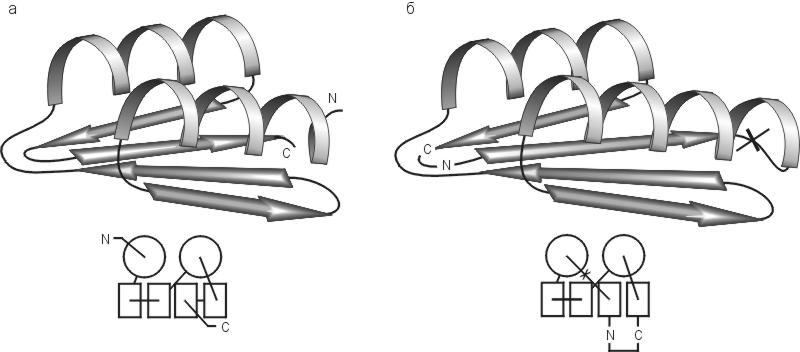
Рис.19-17. Конструируемая пространственная структура альбебетина (а) и схема экспериментально определенной пространственной структуры рибосомального белка S6 (б). Внизу показаны схемы топологий этих белков. Для S6 показана также искусственно введенная пермутация — разрезание петли (![]() ) и соединение исходных N - и С-концов ( N—C ). Такая пермутация придает ему архитектуру, сконструированную для альбебетина.
) и соединение исходных N - и С-концов ( N—C ). Такая пермутация придает ему архитектуру, сконструированную для альбебетина.
Структурное исследование альбебетина показало, что он обладает заданной вторичной структурой (Рис.19-18а) и компактной, весьма стабильной к разворачиванию мочевиной и к протеолизу пространственной структурой. Однако он кооперативно не плавится и находится скорее в состоянии расплавленной глобулы, чем в твердом.
а б 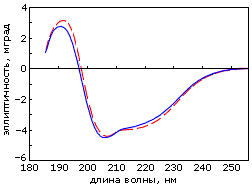
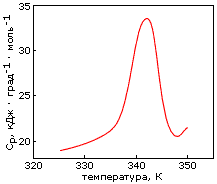
Рис.19-18. (а) Спектр КД альбебетина (![]() ) и альбеферона (
) и альбеферона (![]() ); (б) кривая микрокалориметрического плавления пермутированного, с целью придания ему топологии альбебетина, белка S6. Картинки взяты из D. A.Dolgikh, A. E.Gabrielian, V. N.Uversky & M. P.Kirpichnikov, Appl. Biochem. & Biotech. (1996) 61:85-96, и из Z. Kh. Abdullaev, R. F.Latypov, A. Ya. Badretdinov, D. A.Dolgikh, A. V.Finkelstein, V. N.Uversky & M. P.Kirpichnikov, FEBS Letters (1997) 414:243-246, соответственно.
); (б) кривая микрокалориметрического плавления пермутированного, с целью придания ему топологии альбебетина, белка S6. Картинки взяты из D. A.Dolgikh, A. E.Gabrielian, V. N.Uversky & M. P.Kirpichnikov, Appl. Biochem. & Biotech. (1996) 61:85-96, и из Z. Kh. Abdullaev, R. F.Latypov, A. Ya. Badretdinov, D. A.Dolgikh, A. V.Finkelstein, V. N.Uversky & M. P.Kirpichnikov, FEBS Letters (1997) 414:243-246, соответственно.
Еще один белок со структурой, запланированной для альбебетина, был получен другим методом, — при помощи циркулярной пермутации природного белка S6 (он, как и ряд других белков с недавно расшифрованной пространственной структурой, имеет ту же укладку структурных сегментов, что была разработана для альбебетина, но эти сегменты по-иному соединены белковой цепью, см. Рис.19-17б). Полученный белок обладает твердой, кооперативно плавящейся пространственной структурой (Рис.19-18б).
Недавно альбебетин был использован в качестве носителя функциональной активности. В него — точнее, в новый белок, названый альбефероном — был включен фрагмент 131-138 цепи интерферона a2 человека, способный активировать бласт-трансформацию тимоцитов. Собственно говоря, активацию делает именно это фрагмент интерферона, а остальная, уложенная в глобулу цепь этого белка служит как бы ножнами, — она защищает фрагмент 131-138 от раскусывания и не дает ему действовать слишком уж активно. Опыты показали, что так же работает и альбеферон.
Работы по функционально-активным искусственным белкам ведутся во многих группах. Создаются модели фибриллярных белков на основе длинных спиральных пучков. Создана первая "работающая" модель мембранного белка из амфифильных (гидрофобно-гидрофильных) спиралей, — эти спирали образуют ионные каналы, причем точечные мутации способны резко менять их селективность. Дали функциональный "белок" и a-спиральные полипептиды, химически пришитые к гему.
В общем, в настоящее время белки стремительно превращаются из объекта почтительного и изумленного наблюдения в предмет активной инженерной деятельности (но изумление остается...).
Лекция 20
Сегодняшнюю, заключительную лекцию я посвящу функционированию белков.
Об этом можно говорить очень много, так что мой рассказ даст вам только несколько картинок из жизни работающих белков — картинок, подчеркивающих значение пространственной организации белков для их функции. Некоторые картинки такого рода уже встречались в предыдущих лекциях, — в частности, когда я говорил о мембранных белках. Сегодня я буду говорить только о белках глобулярных, водорастворимых.
Очень грубо, общую схему функционирования белка можно представить в следующем виде:
СВЯЗАТЬ ® ТРАНСФОРМИРОВАТЬ ® ОТПУСТИТЬ
Уточню: некоторые белки делают только часть из этих действий; слова "СВЯЗАТЬ" и "ОТПУСТИТЬ" могут подразумевать связывание и отпуск нескольких разных молекул; а слово "ТРАНСФОРМИРОВАТЬ" может означать и химическую трансформацию, и изменение конформации (как самого белка, так и субстрата), и/или перемещение (белка или субстрата) в пространстве.
Начнем с белков, чья основная функция — "СВЯЗАТЬ". К ним относятся, например, ДНК-связывающие белки.
а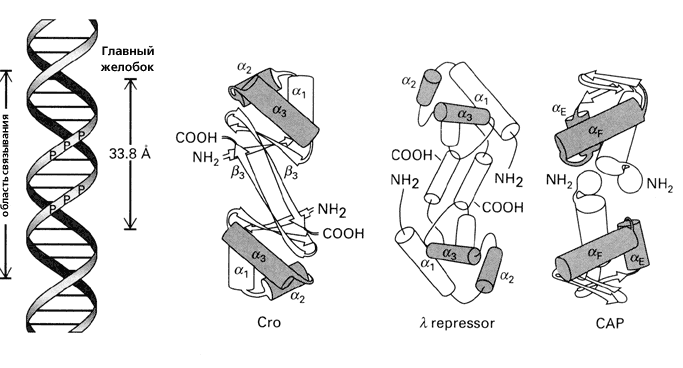 б
б
Рис.20-1 (а) Структура ДНК (слева) и ряда белков, обладающих характерным ДНК-связывающим мотивом "спираль-изгиб-спираль" (он выделен серым цветом). Для белка — активатора катаболитического гена (САР — catabolite gene activator protein) показан только его С-концевой домен. Все эти белки димерны, и все они опознают большой желобок ДНК своими спиралями a3 (aF у САР), расстояние между которыми в димере близко к периоду двойной спирали ДНК (33.8 ![]() ). Картинки взяты из [6]. (б) Изгибание ДНК связавшимся с ней димерным САР-белком (справа, черный). Связывание его с ДНК требует присутствия циклоАМФ (cAMP).
). Картинки взяты из [6]. (б) Изгибание ДНК связавшимся с ней димерным САР-белком (справа, черный). Связывание его с ДНК требует присутствия циклоАМФ (cAMP).
Для связывания с ДНК, поверхность белка — причем на большом протяжении — должна быть приблизительно комплементарна поверхности двойной спирали (Рис.20-1а). Тогда выступы белковой поверхности смогут глубоко внедриться в желобок ДНК, и уже там его боковые группы смогут провести тонкое опознание конкретной ДНКовой последовательности (Рис.20-2) и связаться с той, для которой этот белок предназначен. Все показанные белки на Рис.20-1 — димеры, и именно в такой форме они комплементарны ДНКовому дуплексу. При этом две одинаковые ДНК-опознающие a-спирали такого димера узнают палиндром в двойной спирали ДНК — т. е. такую пару участков в ДНКовом дуплексе, которая одинаково выглядит при перевороте на 180о относительно перпендикулярной к ДНК оси, например
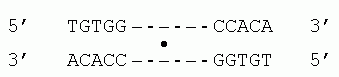
Здесь "-" означают произвольную последовательность ДНК между двумя половинами палиндрома, а "·" — ось переворота.
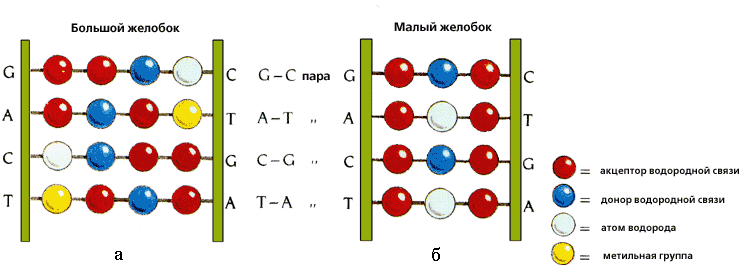
Рис.20-2. Характерные узоры, создаваемые разными функциональным группами A — T и C — G пар в большом и малом желобках ДНК. Картинка взята из [5].
ДНК-связывающие a-спирали в таком белковом димере антипараллельны друг дружке, а расстояние между ними близко к периоду двойной спирали ДНК, так что димер садится на один бок двойной спирали ДНК. Однако разный угол наклона этих a-спиралей к соединяющей их центры оси приводит к тому, что все эти белки по-разному изгибают ДНК при связывании. Порой такой изгиб, индуцированный связыванием, довольно велик (Рис.20-1б).
Иногда связывание белка с ДНК требует наличия кофакторов, которые изменяют — точнее, слегка деформируют — структуру белка и переводят его из неактивной формы в активную.
Так, в trp-репрессоре (он — в E. coli — репрессирует оперон, отвечающий за синтез РНК, кодирующей белки, необходимые для синтеза триптофана), — в trp-репрессоре таким кофатором (точнее, корепрессором) является сам триптофан (Рис.20-3). Пока триптофан не связался с белком, расстояние между ДНК-связывающими спиралями в димере trp-репрессора слишком мало (около 28 ![]() вместо необходимых 34
вместо необходимых 34 ![]() ), так что тот не может связать ДНК. Триптофан же, связавшись с белком, отодвигает спирали так, что они становятся комплементарными к желобку в двойной спирали, — и связываются с ней. Так что когда триптофана в клетке много, — он, связываясь с белком, блокирует дальнейший синтез триптофан-синтезирующих белков, и тем самым — свой (триптофана) дальнейший синтез. Этот способ регуляции называется отрицательной обратной связью.
), так что тот не может связать ДНК. Триптофан же, связавшись с белком, отодвигает спирали так, что они становятся комплементарными к желобку в двойной спирали, — и связываются с ней. Так что когда триптофана в клетке много, — он, связываясь с белком, блокирует дальнейший синтез триптофан-синтезирующих белков, и тем самым — свой (триптофана) дальнейший синтез. Этот способ регуляции называется отрицательной обратной связью.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
