
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
(E2 — E1) = T*(S2 — S1) . | (8.10) |
При небольшом (на dТ) сдвиге температуры от T* свободные энергии фаз немного меняются, так что разность свободных энергий фаз составляет dF = F1(T*+dT) - F2(T*+dT) = dT (S2 - S1). Фазы сосуществуют (т. е. они примерно равновероятны — скажем, соотношение их вероятностей меняется от 10:1 в пользу первой до 1:10 в пользу второй фазы), пока ехр(-dF/k BT*) лежит между 10 и 1/10, т. е. пока dF/kBT* находится где-то от -2 до +2. В этой области |dT| < 2kBT*/(S2-S1).Значит, температурный интервал сосуществования фаз есть
DT » [2kBT*/(S2-S1)] - [-2kBT*/(S2-S1)] = 4kBT*/(S2-S1) = 4T*[kBT*/(E2-E1)] | (8.11) |
Рассмотрим поучительный численный пример. Если T*~300оК (т. е. kBT*~0.6 ккал/моль), а E2-E1 составляет, как то характерно для плавления белков, порядка 50 ккал/моль [т. е. 50/(6x1023) ~ 10-22 килокалорий_на_штуку], то DT — порядка десяти градусов. Если же E2-E1 составляет порядка 50 килокалорий_на_штуку (как при плавлении льдины размером с бутылку), то DT — порядка 10-23 градуса.
Иными словами, для фазовых переходов типа "все-или-ничего" в малых системах и — в гораздо большей степени — для фазовых переходов первого рода в макроскопических системах, — характерен резкий скачок энергии в узком диапазоне температур. В макроскопических системах этот диапазон практически бесконечно узок, а для макромолекул он охватывает несколько градусов, т. е. тоже узок по сравнению с "диапазоном наблюдения", обычно охватывающим от 0оС до 100оС. А вот в небольших олигопептидах резкого скачка нет — здесь диапазон изменения энергии может охватывать все "экспериментальное окошко" исследуемых температур.
Несколько слов о фазовых переходах второго рода. Если для переходов первого рода характерен скачок энергии системы (и ее энтропии, объема и плотности), то для переходов второго рода характерен лишь излом зависимости скорости изменения энергии с температурой, — иными словами, скачок теплоемкости.
Подчеркиваю, что переход второго рода происходит в точке излома (Рис.8-6), а вовсе не посередине часто идущей за ним более или менее "S-образной" зависимости энергии (или другого наблюдаемого параметра) от температуры.
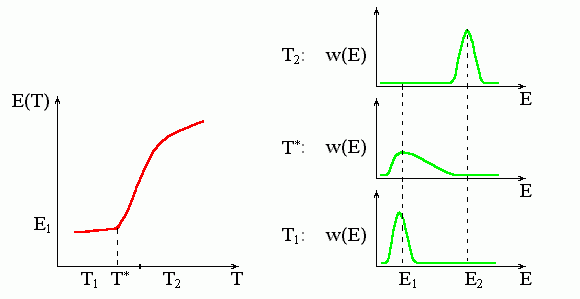
Рис.8-6. Характерная зависимость энергии, E(T), и функции распределения, w(E), при фазовом переходе второго рода. T* — температура этого фазового перехода. При этой температуре функция распределения молекул по энергиям, w(E), резко уширяется, и энергия начинает (или кончает — смотря откуда смотреть) резко изменяться. Обратите внимание, что температура фазового перехода второго рода T* совпадает именно с началом (а не с серединой) резкого изменения энергии.
На рисунках 8-4 — 8-6 я специально представил случаи, когда энергия системы сначала меняется медленно с изменением температуры, потом — резко, а потом — снова медленно. Как вы видите, такое "S-образное" поведение Е(Т) совместимо и с фазовым переходом первого рода, и с градуальным изменением системы при отсутствии какого-либо фазового перехода вообще, и с градуальным изменением, начало которому кладет фазовый переход второго рода.
Я прошу обратить на это особое внимание, так как среди не-физиков существуют два непонятно откуда взявшихся предрассудка: что "переход" соответствует именно середине всякого S-образного изменения, и что если этот "переход" не первого рода — значит второго. Это не так!
Обратите внимание: хотя кривые Е(Т) на рисунках 8-4 — 8-6 выглядят, в общем, одинаково, — поведение кривых распределения по энергиям, w(Е), при переходе первого рода (т. е. типа "все-или - ничего") выглядит совсем не так, как при всех остальных переходах. Для переходов типа "все-или-ничего" — и только для них — характерно наличие двух пиков у w(Е), т. е. сосуществование двух фаз. Поэтому для выяснения того, как — прыжком или постепенно — происходит переход между двумя крайними состояниями, — недостаточно увидеть, что энергия или какой-то наблюдаемый параметр резко меняется в узком диапазоне температур. Здесь нужны дополнительные измерения, которые мы рассмотрим в одной из дальнейших лекций.
В заключение поговорим о кинетике конформационных превращений. Точнее — о том, почему некоторые превращения идут очень медленно. Что здесь значит "медленно"? Предположим, вы знаете время элементарного шага по ходу процесса. Например: один остаток включается во вторичную структуру за 1 наносекунду (нс). Вы знаете также, что в цепи — 1000 звеньев. А процесс идет не за 1000 нс, а за 1000 секунд. Это и есть "медленно": на порядки медленнее, чем можно ожидать, зная время элементарного шага и число таких шагов. Надо понять — почему...
"Медленность" процессов порой связана с медленностью диффузии при высокой вязкости. Однако часто — подчеркиваю, не всегда, но часто — медленность процесса связана с преодолением высокого свободно-энергетического барьера. Особенно это типично для переходов типа "все или ничего", где свободно-энергетический барьер разделяет две фазы (Рис.8-5); здесь он всегда значителен. Такой барьер очень похож на активационный барьер в химических реакциях, — только в данном случае он имеет и энергетическую, и энтропийную составляющие. Напомню, как оценивается скорость такой "барьерной" реакции согласно классической теории переходных состояний.
Рассмотрим простейший процесс, по ходу которого система переходит из состояния А в состояние В. Пусть на пути процесса "А®В" есть один "барьер" # (Рис.8-7), и ни на этом пути, ни "сбоку" от него нет никаких "ловушек" (Рис.8-8: состояний Л, более стабильных, чем исходное A, но менее стабильных, чем конечное B; наличие ловушек усложняет процесс, но не меняет его принципиально).
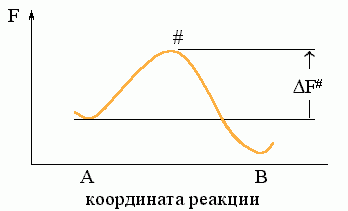
Рис.8-7. Преодоление свободно-энергетического ("активационного") барьера # при переходе из состояния "А" в состояние "В". DF# — свободная энергия барьера (перехoдного состояния).
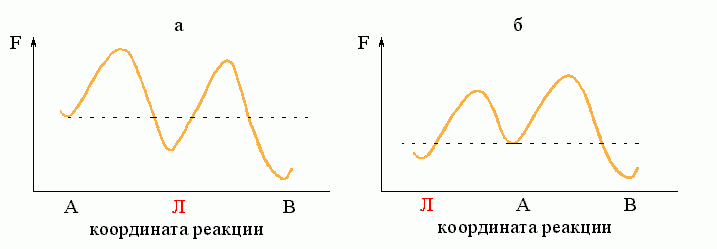
Рис.8-8. "Ловушка": ею может быть как интермедиат "Л" (а), так и побочное (для реакции А®В) состояние "Л" (б). Для наличия кинетической "ловушки" важно только, что она более стабильна, чем исходное состояние "А", но менее стабильна, чем конечное состояние "В", и что на пути из "Л" в "В" находится более высокий свободно-энергетический барьер, чем на пути из А в Л.
Тогда скорость процесса определяется (1) населенностью барьерного ("перех![]() дного") состояния, и (2) скоростью перехода из барьерного в "за-барьерное" состояние.
дного") состояния, и (2) скоростью перехода из барьерного в "за-барьерное" состояние.
Если DF# — свободная энергия барьера, отсчитанная от свободной энергии начального состояния, причем DF# >> kBT, и "ловушек" (Рис.8-8) нет, а в начальном состоянии находится n "частиц" (синонимы: молекул, систем и т. д.), — то на барьере, в каждый момент времени, находится n# ~ n. exp(-DF#/kBT) частиц. Пусть каждая из этих на-барьерных частиц переходит за барьер за время t (время элементарного шага реакции). Тогда за время порядка t за барьер перейдут все n# на-барьерных частиц. На то, чтобы за барьер перевалили бы все n частиц — понадобится n/n# шагов, т. е. время порядка
t ~ t (n/n#) ~ t exp(+DF#/kBT) | (8.12) |
При наличии "ловушки", Л, таким же образом оценивается время перехода из А в Л и из Л в В.
Уточнение. Если на пути из А в В находится не одно, а s последовательных промежуточных состояний с со свободной энергией, близкой к DF#, — то скорость процесса замедляется в s2 раз по сравнению с оценкой (8.12). Это замедление связано с диффузией частиц по верхушке барьера. Однако обычно этот диффузионный фактор не играет существенной роли.
Лекция 9
Обсудим теперь, зная базовую физику, стабильности вторичной структуры и кинетику ее образования.
Начнем с a-спиралей. Первая водородная связь в ней, (СО)0 — (HN)4, фиксирует конформации трех остатков — 1, 2, 3; следующая водородная связь, (СО)1 — (HN)5, дополнительно фиксирует конформацию только одного остатка — остатка 4; связь (СО)2 — (HN)6 дополнительно фиксирует остаток 5, и т. д.
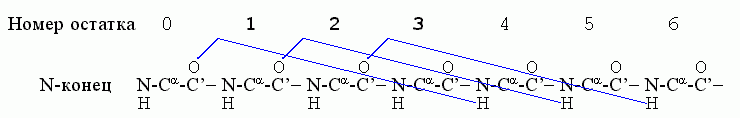
Значит, если в спирали фиксировано n остатков, то их фиксирует n-2 водородные связи. Рассмотрим свободную энергию образования такой спирали из клубка в водном окружении ("клубок" — это полимер без структуры и без взаимодействия дальних по цепи звеньев). Эту свободную энергию можно записать как
DFa = Fa — Fклуб. = (n-2)fH — nTSa = -2fH + n(fH —TSa) | (9.1) |
Здесь fH — свободная энергия водородной связи в a-спирали (вы помните, что fH — не просто энергия, как было бы в вакууме: в нее входит и энергия, и энтропия перестроек водородных связей в водном окружении), а Sa — потеря энтропии при фиксации одного остатка в спирали.
Вы видите, что в DFa есть два члена. Один (-2fH) не зависит от длины спирали; величина

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
