
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
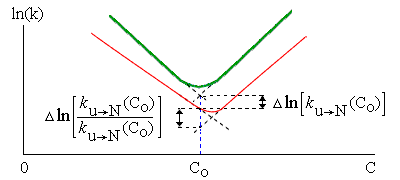
Рис.17-9. Схема, иллюстрирующая сдвиг "шевронного графика" при мутации. Жирная линия: исходный белок; тонкая — мутант. Пунктир показывает экстраполяцию величин ku®N и kN®u в области излома шеврона. С0 — концентрация денатуранта, соответствующая середине денатурационного перехода в исходном белке. На графике показаны измеряемые в этом опыте величины. Одна из них определяет влияние мутации на высоту свободно-энергетического барьера, стоящего на пути из развернутого состояния в нативное: D(F#-Fu) = -RTDln[ku®N]. Вторая — ее влияние на стабильность белка, т. е. на разность свободных энергий нативного и денатурированного состояний: D(FN-Fu) = -RTDln[ku®N/kN®u]. Показанные на графике величины относятся к концентрации денатуранта C0, где требующаяся экстраполяция минимальна (а потому минимальны и погрешности в ней), — но, ценой несколько большей экстраполяции (до C0=0), так обычно определяются изменения в стабильности нативного белка, D(FN-Fu), и в высоте барьера, D(F#-Fu), относящиеся к чистой воде.
Для оценки вовлеченности остатка в переходное состояние (или, как говорят, в "зародыш" сворачивания белка), оценивают, по сдвигу шеврона, влияние мутации данного остатка на (а) скорость сворачивания белка и (б) на его стабильность. Измеряемые величины показаны на Рис.17-9.
Скорость сворачивания белка (перехода U®N) определяется (см. формулу 17.1) разностью свободной энергии "зародыша" (#) и исходного развернутого (U) состояния белка (т. е. величиной F#-Fu).
Если мутация остатка так же влияет на величину F#-Fu, как она влияет на стабильность всего нативного состояния (т. е. на величину FN-Fu), — то это свидетельствует о том, что рассматриваемый остаток так же вовлечен в "зародыш" (образует там те же контакты, имеет ту же конформацию и т. д.), как он вовлечен в нативную глобулу.
Если, наоборот, мутация остатка оказывает влияние только на стабильность белка (т. е. на величину FN-Fu), но не на скорость сворачивания (т. е. она не влияет на величину F#-Fu), — значит, этот остаток не вовлекается в глобулярный "зародыш" сворачивания, т. е. что он входит в нативную белковую глобулу только уже после образования зародыша.
И, наконец, если мутация остатка сильно влияет на стабильность белка и слабее (но с тем же знаком) — на стабильность зародыша, — значит, этот остаток образует в зародыше только часть тех контактов, которые имеет в нативном белке.
Так очерчивается зародыш сворачивания белка (Рис.17-10): для каждого из мутированных остатков цепи вычисляется величина
Ff = D(F#-Fu)/D(FN-Fu) , — | (17.8) |
и, если для данного остатка Ff близка к 1, — то говорят, что он входит в зародыш структуры; а если Ff близка к 0 — то нет.
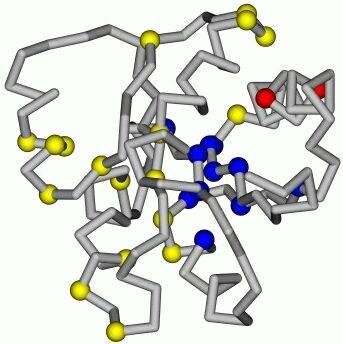
Рис.17-10. Структура переходного состояния белка CheY, согласно T. L![]() pez-Hern
pez-Hern![]() ndes & L. Serrano, Folding & Design (1996) 1:43-55. На фоне нативной укладки цепи CheY синими шариками выделены остатки, вовлеченные в переходное состояние (образующие там более 30% своих контактов), желтыми шариками — не вовлеченные в переходное состояние остатки. Без шариков оставлены те области цепи, где мутации еще не делались. Красным закрашены остатки цепи, сложные для интерпретации, — у них столь малы величины D(F#-Fu) и D(FN-Fu) (последнее важнее, так как эта величина стоит в знаменателе в формуле 17.8), что погрешности в их определении превышают сами эти величины.
ndes & L. Serrano, Folding & Design (1996) 1:43-55. На фоне нативной укладки цепи CheY синими шариками выделены остатки, вовлеченные в переходное состояние (образующие там более 30% своих контактов), желтыми шариками — не вовлеченные в переходное состояние остатки. Без шариков оставлены те области цепи, где мутации еще не делались. Красным закрашены остатки цепи, сложные для интерпретации, — у них столь малы величины D(F#-Fu) и D(FN-Fu) (последнее важнее, так как эта величина стоит в знаменателе в формуле 17.8), что погрешности в их определении превышают сами эти величины.
Замечательно, что лишь очень малое число остатков в белке не удается проинтерпретировать с такой точки зрения (а с нее не удалось бы проинтерпретировать такие остатки, чьи Ff величины лежали бы вне интервала 0 — 1, т. е. те остатки, мутации которых влияли бы только на скорость сворачивания, но не на стабильность нативной структуры; или те, что стабилизировали бы только зародыш, но дестабилизировали бы нативный белок, и т. д.).
Это вселяет уверенность в том, что базовая картина, согласно которой остатки — уж если они вовлечены в зародыш — стоят там так же, как в нативном белке, — эта картина в основном справедлива.
Рисунок 17-10 показывает, что остатки, наиболее существенные для сворачивания белка, группируются в компактный "доменчик", компактное ядрышко сворачивания, причем оно лежит не в геометрическом центре белка, а сдвинуто к его поверхности. Аналогичная картина наблюдается и в других (впрочем, пока немногочисленных) белках, исследованных на предмет местоположения их ядер сворачивания.
В заключение я хочу вернуться к Рис.17-8 и подчеркнуть, что даже для совершенно конкретного белка не существует четко определенного "характерного времени сворачивания". Действительно, этот рисунок показывает, что сворачивание лизоцима занимает порядка 0.1 сек в нативных условиях, и порядка 10000 сек — в условиях созданного гуанидингидрохлоридом (при той же температуре раствора) равновесия нативной и денатурированной форм. А в других условиях, при повышенной температуре, но без гуанидингидрохлорида, области равновесия этих форм отвечает (см. Рис.17-6) время сворачивания порядка 10 сек. Так что, обсуждая скорость сворачивания белка, мы должны иметь в виду либо весь наблюдаемый диапазон времен сворачивания данного белка, либо совершенно конкретные экспериментальные условия — например, те условия, при которых белок сворачивается в клетке.
Лекция 18
Сегодня мы продолжим разговор о самоорганизации белков.
Все экспериментальные данные, о которых я рассказывал, — сколь бы интересны они ни были сами по себе — не дают ответа на вопрос, как белок ухитряется найти свою нативную структуру — среди астрономического числа возможных! — за те немногие секунды или доли секунды, что отпущены на его сворачивание.
А число это — как я говорил, его оценил в еще 1968 г. Сайрус Левинталь — действительно огромно: ~10100 возможных конформаций для цепи из 100 остатков; их "тупой" перебор занял бы ~1080 лет — кладя всего 10-13 секунды на переход из одной конформации в другую.
Как же белок выбирает свою нативную структуру среди бесчисленного множества возможных? — спросил Левинталь, и ответил: — По-видимому, самоорганизующийся белок следует по какому-то специальному "пути сворачивания", и та структура, где этот путь заканчивается, и является его нативной структурой. Иными словами, Левинталь предположил, что нативная структура белка определяется не стабильностью, не термодинамикой, а кинетикой, т. е. она соответствует не глобальному, а просто быстро достижимому минимуму свободной энергии цепи.
Сложность проблемы заключается в том, что поднятый вопрос нельзя решить чисто экспериментально. Действительно: предположим, у белковой цепи есть другая, "ненанивная", но еще более стабильная укладка. Как ее найти, если сам белок ее не находит? Ждать результата в течение 1080 лет?!
С другой стороны, вопрос о том, что — кинетика или термодинамика — определяет укладку белковой цепи, постоянно возникает на пути решения разных прикладных задач. Он возникает, когда речь идет о предсказании структуры белка по его аминокислотной последовательности (надо знать, что предсказывать: самую стабильную или самую быстро сворачивающуюся его структуру). Он возникает и тогда, когда речь идет о дизайне новых, не встречающихся в природе белков (надо знать, что делать: максимально усиливать стабильность желаемой структуры или пролагать максимально быстрый путь к ней).
Однако действительно ли существует противоречие между "структурой стабильной" и "структурой быстро сворачивающейся"? Может быть, стабильная структура автоматически обладает свойством сворачиваться быстро?
Прежде, чем приступить к исследованию этих вопросов, т. е. прежде чем рассматривать кинетические аспекты сворачивания белков, вспомним ряд фундаментальных фактов из области их термодинамики (здесь всюду речь идет об относительно небольших, однодоменных белках, т. е. о белках из 50—200 аминокислотных остатков). Эти факты помогут нам понять, какие условия протекания процесса сворачивания мы должны рассматривать. Термодинамические факты таковы:
1) Разворачивание белка обратимо, причем оно происходит как переход "все-или-ничего". Последнее означает, что в точке денатурации белка только две формы белковой молекулы — "нативная" и "денатурированная" — присутствуют в заметных количествах, а все прочие ("полусвернутые" и "неверно свернутые" формы) практически отсутствуют.
2) Денатурированная форма белков — во всяком случае, небольших белков, развернутых денатурантом, — часто является неупорядоченным клубком.
3) В нормальных физиологических условиях нативная форма белка лишь немногим стабильнее его развернутой формы (а в сам![]() й точке плавления обе эти формы имеют, естественно, одинаковую стабильность). При этом нативная структура белка стабильна благодаря своей низкой энергии, т. е. благодаря сильным взаимодействиям в нативной структуре, а развернутая, — благодаря своей высокой конформационной энтропии, т. е. благодаря огромному числу разных развернутых конформаций. [Необходимое пояснение: как принято в литературе, термин "энергия" здесь означает, строго говоря, всю свободную энергию взаимодействий, в том числе взаимодействий цепи с растворителем (например, "энергия" гидрофобных взаимодействий определяется, как вы должны помнить, энтропией растворителя); термин же "энтропия" здесь охватывает лишь конформационную энтропию цепи, но не энтропию растворителя. Такая терминология принята, чтобы, оставив растворитель за скобками, сосредоточиться на главной проблеме, — как белковая цепь находит "свою" пространственную структуру среди гигантского числа возможных.]
й точке плавления обе эти формы имеют, естественно, одинаковую стабильность). При этом нативная структура белка стабильна благодаря своей низкой энергии, т. е. благодаря сильным взаимодействиям в нативной структуре, а развернутая, — благодаря своей высокой конформационной энтропии, т. е. благодаря огромному числу разных развернутых конформаций. [Необходимое пояснение: как принято в литературе, термин "энергия" здесь означает, строго говоря, всю свободную энергию взаимодействий, в том числе взаимодействий цепи с растворителем (например, "энергия" гидрофобных взаимодействий определяется, как вы должны помнить, энтропией растворителя); термин же "энтропия" здесь охватывает лишь конформационную энтропию цепи, но не энтропию растворителя. Такая терминология принята, чтобы, оставив растворитель за скобками, сосредоточиться на главной проблеме, — как белковая цепь находит "свою" пространственную структуру среди гигантского числа возможных.]

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
