
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Чем же плохи перекрывания петель — ведь при этом цепи не врезаются друг в друга, а просто одна из них проходит на другой? Тем, что "нижняя", прижатая к ядру петля лишается водородных связей с водой. А чтобы залечить эту потерю — опять нужны "редкие" последовательности...
Здесь, правда, нас должно смутить то, что при перекрывании петель пропадет лишь одна, максимум две водородные связи — т. е. энергии потеряется немного, килокалорий три или пять. Это не только много меньше, чем полная энергия взаимодействий в белке, измеряемая обычно сотнями килокалорий (судя по опытам по плавлению белков), но и заметно меньше чем обычный "запас стабильности" белка (т. е. разность свободных энергий его нативного и денатурированного состояния). В нативных условиях этот запас составляет — по тем же опытам — порядка 10 ккал/моль. Почему же "дефект" ценой всего в 5 ккал/моль запрещает — или почти запрещает — перекрывание петель в нативных белковых глобулах?
И еще вопрос: что мешает сделать в петле дополнительный изгиб (пунктир на Рис.15-4) — и тем избежать физического перекрывания одной петли другой (т. е. заменить перекрывание петель обходом)? Может быть, здесь дело в упругости полимерной цепи — ведь за дополнительный изгиб петле пришлось бы заплатить (как показывает расчет) несколько (все те же несколько!) ккал/моль?
Здесь любой человек, знающий физику полимеров, должен прервать меня и сказать: "Упругость полимера — энтропийный, а вовсе не энергетический эффект! То есть сильно изогнутая цепь не может флуктуировать так свободно, как прямая или слабо изогнутая. Иными словами, с сильно изогнутой формой цепи совместимо гораздо меньше конформаций, чем с ее более или менее вытянутой формой. Однако эффект, о котором Вы говорите, относится к флуктуирующей цепи, — т. е. к цепи, не фиксированной в глобуле. Но в нативном белке цепь фиксирована, и — так ли она идет, иначе ли — она все равно будет иметь лишь одну какую-то конформацию. Какое отношение имеют описываемые Вами энтропийные потери к нативной структуре белка, где цепь все равно фиксирована, т. е. все равно имеет нулевую энтропию?"
Запомним эти вопросы, а пока рассмотрим еще одну характерную черту белковых архитектур — то, что перемычка между параллельными b-участками почти всегда образует с ними правозакрученную, а не левозакрученную спираль (Рис.15-5).
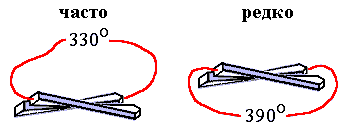
Рис.15-5. Левовинтовой ход перемычек между параллельными b-участками очень редко наблюдается в белках, правовинтовой — часто.
Критерий стабильности позволяет и в этом случае выделить "лучшую" из двух зеркально-симметричных белковых архитектур. В основе различия лежит зеркальная асимметрия природных аминокислот. Она приводит, как вы помните, к преимущественно правому, если смотреть по ходу b-тяжей, скручиванию b-слоев, состоящих из L-аминокислот (это скручивание показано на Рис.15-5). При этим угол между осями соседних b-участков близок к 300, так что полный угол поворота близок к 3300 для правовинтовой перемычки и к 3900 — для левовинтовой. В результате, из-за жесткости полипептидной цепи, правовинтовая — менее закрученная — перемычка выгоднее, чем левовинтовая, т. е. ее свободная энергия ниже, — хотя и опять немного, на пару ккал/моль, и опять в результате не энергетического, а энтропийного эффекта.
И здесь мы опять наталкиваемся на два поставленных выше (и пока оставленных без ответа) вопроса:
(1) Почему "дефект" ценой всего в несколько ккал/моль — на фоне гораздо большей полной энергии белка — может практически запрещать многие мотивы белковых архитектур?
(2) Какое отношение имеют описываемые здесь упругие, т. е. энтропийные эффекты к нативной структуре белка, где цепь все равно фиксирована?
Начнем с первого вопроса — вопроса о проявлении энергии "дефекта" в статистике белковых архитектур. Но сначала усугубим его. Посмотрим, как связаны с энергией другие статистические закономерности, отмеченные в белковых структурах? Оказывается — точно так же! Но здесь мы располагаем большей статистикой и можем получить не только качественные, но и количественные оценки.
Для примера рассмотрим статистику распределения аминокислотных остатков между внутренностью и поверхностью белковой глобулы и посмотрим, как она связана с гидрофобностью аминокислотных остатков. Гидрофобность аминокислотных остатков обычно измеряется в свободной энергии их переноса из октанола, моделирующего гидрофобное ядро белка, в воду. На Рис.15-6 эта гидрофобность отложена по вертикальной шкале. По горизонтальной шкале отложен логарифм отношения числа поверхностных и внутренних остатков в белках — точнее, этот логарифм, умноженный на RT, где Т=3000К соответствует примерно комнатной температуре. Мы видим, что точки более или менее ложатся на прямую линию — и наклон такой прямой более или менее близок к 1 — 1.5.
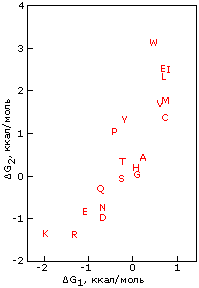
Рис.15-6. Экспериментально определенная свободная энергия переноса боковых групп аминокислотных остатков из неполярного растворителя в воду (DG2), и "кажущаяся свободная энергия переноса остатка из ядра на поверхность белка" (DG1), вычисленная из наблюдаемых частот встречаемости аминокислотных остатков внутри (fin) и на поверхности (fsurf) белка по формуле DG1 = - RT ln[fsurf/fin]. Картинка взята из S. Miller, J. Janin, A. M.Lesk, C. Chothia, J. Mol. Biol. (1987) 196:641-656.
Таким образом, наблюдаемая статистика распределения остатков между нутром и поверхностью глобулы неплохо аппроксимируется формулой
ВСТРЕЧАЕМОСТЬ ~ exp(-СВОБОДНАЯ_ЭНЕРГИЯ/kTC) , | (15.1) |
где ТС — какая-то температура, близкая не то к комнатной температуре, не то к характерной температуре плавления белка: ведь 3000К и 3500К совпадают "по порядку величины".
То есть статистка встречаемости аминокислотных остатков внутри и на поверхности белка удивительно похожа на статистику Больцмана по форме! Это впервые было замечено Полем в 1971 г. для распределения углов внутреннего вращения в боковых цепях аминокислотных остатков в белках. Потом это показано и для статистики многих других элементов белковых структур: для встречаемости ионных пар, для встречаемости остатков во вторичных структурах, для встречаемости полостей в белках и т. д., и т. п. К настоящему времени эта аналогия стала столь привычной, что статистика белковых структур часто используется для оценки свободной энергии различных взаимодействий аминокислотных остатков.
Здесь, однако, следует подчеркнуть, что белковая статистка похожа на статистику Больцмана именно по своей экспоненциальной форме, а не по физическому смыслу. Напомню, что статистика Больцмана поддерживается тем, что частицы бродят с места на место, и каждая из них больше времени проводит там, где ее энергия ниже. В то же время в нативных белках аминокислотные остатки не бродят с места на место! Например, Leu72 цепи миоглобина кашалота всегда находится внутри глобулы в нативной структуре этого белка, — мы никогда не видим его на поверхности. И если мы видим, по статистике, что 80-85% всех лейцинов находится внутри белка и 15-20% на поверхности, — то здесь дело не в том, что каждый лейцин проводит 80-85% времени внутри и 15-20% времени на поверхности глобулы. Здесь дело в том, что естественный отбор закрепил большинство лейцинов в тех точках цепи, что лежат внутри глобулы.
То есть обычная Больцмановская статистика здесь, в распределении остатков между ядром и поверхностью белка — вовсе ни при чем.
Посмотрим с другой стороны.
Чем хорошо, скажем, преимущественное расположение лейцинов внутри глобулы? — Тем, что оно повышает ее стабильность. Но почему тогда естественный отбор не настоял на том, чтобы все лейцины были бы внутри белка? Может быть, потому, что уже 80-85% внутренних лейцинов достаточно для стабильности белка, а возня с остальными потребовала бы от него слишком больших усилий?
Оставим вопрос о психологии естественного отбора как бесперспективный и ненаучный, и рассмотрим другой вопрос — вопрос о том, как изменяет внутренняя свободная энергия какого-то элемента белковой структуры число аминокислотных последовательностей, способных придать стабильность белку с этим структурным элементом. Например — сравним число стабилизирующих данную пространственную структуру последовательностей при условии, что в такой-то лежащей внутри белка точке цепи находится лейцин, с числом стабилизирующих ту же структуру последовательностей при условии, что в этой точке цепи находится серин.
Иными словами — посмотрим, как изменится число стабилизирующих структуру белка последовательностей при мутации Leu ® Ser во внутренней точке белка.
Нативная структура стабильна, если ее свободная энергия меньше, чем свободная энергия денатурированного белка. Будем, для простоты, считать, что (1) вклад остатка в стабилизацию белка определяется свободной энергией его дегидратации [что качественно справедливо, хоть может быть и не совсем верно количественно], (2) что внутренние остатки белка целиком укрыты от воды, а поверхностные - целиком ей доступны [что является довольно грубым, но качественно верным допущением], и что денатурированный белок совсем развернут [это бывает часто, но не всегда; поэтому излагаемая ниже теория приблизительна, — но зато она проста.]
Свободная энергия переноса серина из гидрофобного окружения в воду — около 0, а лейцина — около +2 ккал/моль. В развернутом денатурированном белке все остатки окружены водой. Значит, структура белка с серином во внутренней точке "в среднем" (по разным последовательностям) на 2 ккал/моль менее стабильна, чем структура белка с лейцином в той же внутренней точке. Следовательно, грубо говоря, все последовательности, стабилизирующие белок с серином в ядре, будут стабилизировать и белок с лейцином в ядре, — но, кроме того, белок с лейцином внутри будут стабилизировать и какие-то последовательности, не способные стабилизировать белок с серином в той же точке.
Как упадет число стабилизующих нативную структуру белка последовательностей при замене более стабильного элемента этой структуры ("Leu внутри") на менее стабильный ("Ser внутри")?

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
