
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рис.1-5. Два разных по форме, но почти идентичных по функции белка (сериновые протеазы): химотрипсин и субтилизин. Несмотря на кардинальное различие в укладке цепи, их каталитические центры состоят из одинаковых и одинаково в пространстве расположенных остатков: Ser195 (оранжевый), His57 (синий) и Asp102 (малиновый) в химотрипсине, и Ser221 (оранжевый), His64 (синий) и Asp32 (малиновый) в субтилизине.
Однако общим у этого "всего остального" тела является то, что тело белка — твердое: иначе нельзя обеспечить специфичность активного центра.
В свое время мы подробно рассмотрим структуры белков, их способность к самоорганизации и причину их твердости, обсудим функции и прочие интересные для биолога вопросы, — но сначала необходимо рассмотреть аминокислотные остатки и их элементарные взаимодействия между собой и с окружающей средой.
ЭЛЕМЕНТАРНЫЕ ВЗАИМОДЕЙСТВИЯ В БЕЛКАХ И ВОКРУГ
Лекция 2
Аминокислоты, из которых сложена полипептидная цепь (Рис.2-1), могут находиться в двух стерических формах: L и D.
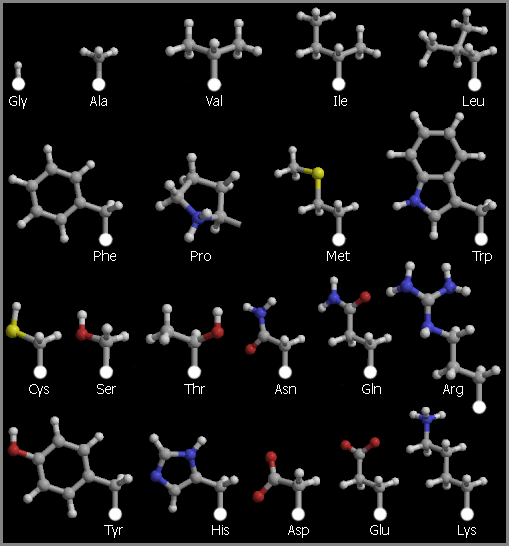
Рис.2-1. Боковые цепи двадцати стандартных аминокислотных остатков, исходящие из Сa атомов главной цепи. Белыми кружками показаны Сa атомы главной цепи, серыми — С атомы боковых групп, светло-серыми — H атомы боковых групп, синими — N атомы боковых групп, красными — O атомы боковых групп, желтыми — S атомы боковых групп.
Эти формы, L и D, зеркально-симметричны: в них массивный боковой радикал (R) и Н-атом, стоящие при a-углероде (Сa) аминокислоты, меняются местами (стрелки указывают углубление атомов в рисунок):
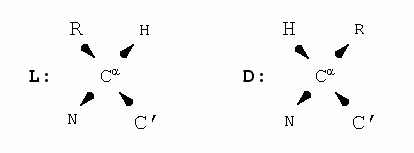
L и D форм нет только у глицина (Gly), "боковая цепь" которого состоит лишь из Н-атома.
Белковые цепи сложены только из остатков L-аминокислот. Только они кодируются генами. D-аминокислотные остатки — они встречаются в пептидах — не кодируются при матричном синтезе белка, а синтезируются специальными ферментами. Рацемизация (L « D переход) спонтанно в белках практически не происходит. Не происходит она и при биосинтезе, — хотя часто встречается при чисто химическом синтезе пептидов, где на борьбу с рацемизацией уходит много сил.
Аминокислоты в белковой цепи связаны между собой пептидными связями C' и N атомов (Рис.2-2).
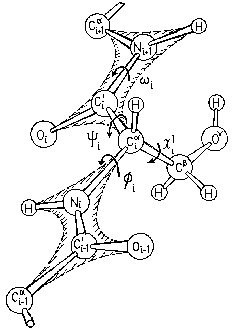
Рис.2-2. Полипептид: главная цепь и боковая группа (Ser) на ней. Пептидные группы заштрихованы. Показаны углы внутреннего вращения в главной (f, y, w) и боковой (c) цепях. Стрелки указывают направление вращения, ведущее к росту угла поворота ближней к нам части цепи относительно более отдаленной ее части. Картинка взята из [3] и адаптирована.
Важную роль в структуре белка играет как жесткость этих связей, так и плоская форма всей пептидной группировки
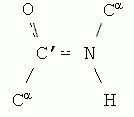
И то, и другое обеспечивается так называемой sp2-гибридизацией электронов N и С' атомов. "Гибридизация" электронных орбит — это чисто квантовый эффект. Sp2-гибридизация преобразует одну сферическую s- и две вытянутые p-орбиты электронов атома в три sp2-орбиты. Эти три орбиты вовлекают атом в три ковалентные связи, лежащие в одной плоскости (-·<). Вовлеченные в sp2-гибридизацию электроны ковалентно связанных C' и N атомов создают "делокализованное" электронное облако, охватывающее оба эти атома, и распространяющиеся также — через связь С-О — на атом О (именно поэтому связи C'![]() N и C
N и C![]() O рисуются как равноправные "полуторные",
O рисуются как равноправные "полуторные", ![]() ). Но у C и N атомов есть еще p-электроны кроме тех, что уже задействованных в sp2 орбитах! И так как эти p-орбиты имеют форму "восьмерок", перпендикулярных всем трем sp2-орбитам (-·<), то дополнительная связь этих p-электронов — связь, требующая сближения "восьмерочных" орбит и "обобществления" (т. е. переходов с атома на атом) находящихся на них электронов,
). Но у C и N атомов есть еще p-электроны кроме тех, что уже задействованных в sp2 орбитах! И так как эти p-орбиты имеют форму "восьмерок", перпендикулярных всем трем sp2-орбитам (-·<), то дополнительная связь этих p-электронов — связь, требующая сближения "восьмерочных" орбит и "обобществления" (т. е. переходов с атома на атом) находящихся на них электронов,
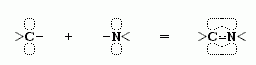
— это обобществление p-электронов препятствует вращению вокруг С'-N связи.
Хочу напомнить, что делокализация электронов, их переход с атома на атом — основная причина образования химических связей. Это следует из принципа неопределенности Гайзенберга:
DpDx » | (2.1) |
Здесь Dp — неопределенность в импульсе частицы, Dx — неопределенность в ее координате, а ![]() º h/2p, где h — постоянная Планка. Так как направление движения электрона в атоме не может быть известно, то Dp » |p| = mv, где v — скорость, а m — масса частицы. Следовательно,
º h/2p, где h — постоянная Планка. Так как направление движения электрона в атоме не может быть известно, то Dp » |p| = mv, где v — скорость, а m — масса частицы. Следовательно,
v » ![]() /(mDx) .
/(mDx) .
В то же время, кинетическая энергия частицы E = mv2/2, т. е.
E » | (2.2) |
Следовательно, при делокализации, когда Dx возрастает, — энергия частицы падает, и она переходит в более стабильное состояние. Видно, что этот эффект наиболее силен для легких частиц — т. е. электронов. Так делокализация электрона приводит к химической связи.
Длина химической связи близка к Вандерваальсову радиусу атома, т. е. составляет 1 — 2![]() (точнее: около 1
(точнее: около 1 ![]() — для связи C-H, N-H или O-H, около 1.2 — 1.3
— для связи C-H, N-H или O-H, около 1.2 — 1.3 ![]() — для С=О, C
— для С=О, C![]() O, C
O, C![]() N и С=С, 1.5
N и С=С, 1.5 ![]() — для С-С, и около 1.8
— для С-С, и около 1.8 ![]() — для S-S).
— для S-S).
Характерные величины валентных углов составляют около 120о и около 109о. Углы в 120о находятся при sp2-гибридизованных атомах типа - C'<, - N<, где три валентные связи направлены из центра к вершинам плоского треугольника, углы в 109о — при sp3-гибридизованных атомах типа >Ca<, где четыре связи направлены из центра к вершинам тетраэдра, а также при завязывающих по две связи O< или S< атомах.
Остановимся теперь на характерных величинах флуктуаций, т. е. тепловых колебаний валентных связей и углов. Они могут вносить вклад в гибкость белковой цепи.
Частоты колебаний проявляются в инфракрасных (ИК) спектрах белков. Характерные частоты колебаний таковы: n ~ 7x1013 сек-1 — для колебания атома Н, — например, в С-Н связи (при этом длина волны l=c/n~5мк); и n ~ 2x1013 сек-1 — для колебания "тяжелых" групп, — например, в СН3-СН3 связи (при этом l=c/n~15мк; здесь c — скорость света, 300000 км/сек).
Возбуждены ли эти колебания теплом при комнатной температуре?
Для ответа на этот вопрос мы должны сравнить тепловую энергию, приходящуюся на одну степень свободы ("тепловой квант" kT) с энергией колебания. Оценим величину kT при "нормальной" температуре. Здесь T — абсолютная температура в градусах Кельвина (Т=300оК при 27оС, т. е. при примерно "комнатной" температуре; К — обозначение градуса Кельвина), а k (иногда еще пишут kB) — постоянная Больцмана [равная 2 (кал/моль)/К, или 0.33x10-23 кал/К в расчете на одну частицу, так как в одном моле — 6x1023 частиц]. Значит, при комнатной температуре "тепловой квант" kT = 600 кал/моль, или (600кал)/(6x1023 частиц), т. е. 10-21 калорий на частицу.
Соответствующую такому тепловому кванту частоту nT можно найти из соотношения kT = hnT [где постоянная Планка hº2p![]() =6.6x10-34 Джxсек = 1.6x10-34 калxсек; напомню, что 1 калория (кал) = 4.2 джоуля (Дж)]. Итак, частота nT равна 7x1012 сек-1 при T=300K, т. е. при 27оС.
=6.6x10-34 Джxсек = 1.6x10-34 калxсек; напомню, что 1 калория (кал) = 4.2 джоуля (Дж)]. Итак, частота nT равна 7x1012 сек-1 при T=300K, т. е. при 27оС.
"Тепловой квант" не может возбудить более жесткие, более частые, чем он сам, колебания.
Значит, при комнатной температуре валентные связи "тверды" и не флуктуируют: для этих связей частота колебаний n ~ 2x1013 — 7x1013 сек-1, т. е. она на порядок больше, чем nT = 7x1012 Джxсек-1.
Однако валентные связи могут раскачиваться падающим инфракрасным светом, и на этом основана ИК-спектроскопия белков. Собственно, именно ИК спектроскопия и дает сведения о колебаниях атомов, валентных связей и валентных углов. Конечно, экспериментально их исследуют на малых молекулах, а затем полученную информацию используют при изучении белков.
В отличие от колебаний длин связей, тепловые колебания валентных углов, на порядок более мягкие, — их частоты лежат в пределах 1012 — 1013 сек-1, т. е. она на порядок больше, чем nT = 7x1012 Джxсек-1.
Однако валентные связи могут раскачиваться падающим инфракрасным светом, и на этом основана ИК-спектроскопия белков. Собственно, именно ИК спектроскопия и дает сведения о колебаниях атомов, валентных связей и валентных углов. Конечно, экспериментально их исследуют на малых молекулах, а затем полученную информацию используют при изучении белков.
В отличие от колебаний длин связей, тепловые колебания валентных углов, на порядок более мягкие, — их частоты лежат в пределах 1012 — 1013 сек-1, — возбуждены при комнатной температуре. Однако их характерная амплитуда невелика — около 5о.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
