
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Теперь давайте более подробно рассмотрим фотосинтетический реакционный центр (Рис.12-5). Его задача — проводить выбитые светом электроны на другую сторону мембраны, и тем самым создавать тот электрический потенциал, на котором и работает фотосинтез.
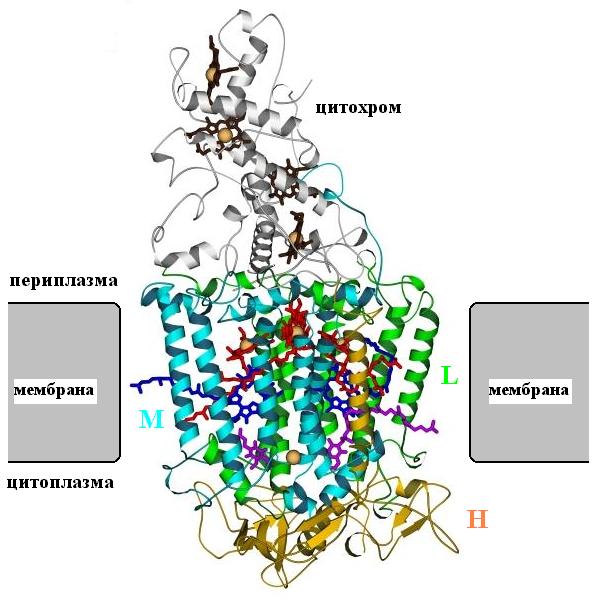
Рис.12-5. Фотосинтетический реакционный центр. Мембрана показана схематично. Трансмембранная субъединица М выделена голубой раскраской, трансмембранная субъединица L — зеленой, субъединица Н (у нее есть одна трансмембранная спиралью) — желтой; цитохром — серый. Обратите внимание, насколко внутримембранные части регулярнее внемембранных! Субъединицы L и М связывают фотосинтетические пигменты — хлорофиллы (красные с желтым ионом магния в центре) и феофитины (темно-синие). И у тех, и у других есть длинные гидрофобные "хвосты", выступающие из белка в мембрану. Субъединицы L, M связывают и два хинона (выделенные фиолетовой окраской). Цитохром, лежащий вне мембраны, включает четыре гема (черно-серые, с оранжевыми ионами железа внутри). Все кофакторы изображены в виде скелетных моделей; см. также Рис.12-6.
Фотосинтетический реакционный центр включает в себя цитохром с четырьмя гемами (это, собственно говоря, — не мембранный белок: он находится вне ее, в периплазматическом пространстве) и три мембранные субъединицы: L, M и H. (от последней, впрочем, в мембране находится только одна a-спираль). Субъединицы L и M очень похожи.
Все трансмембранные части — a-спирали. Они, как всегда, длинны (их длина равна толщине мембраны) и регулярны. Внутри мембраны нет нерегулярных петель. В то же время лежащая вне мембраны цепь значительно менее регулярна и содержит много петель; в общем, она уложена так же, как в "обычных" водорастворимых глобулярных белках, о которых нам еще предстоит говорить.
Обратите внимание на множество небольших циклических молекул, сидящих в белке фотосинтетического реакционного центра: именно по сложенным из них дорожкам, "проводникам" идет электрон (это видно по изменениям электронных спектров этих молекул), а сам полипептид служит только, так сказать, формообразующим изолятором.
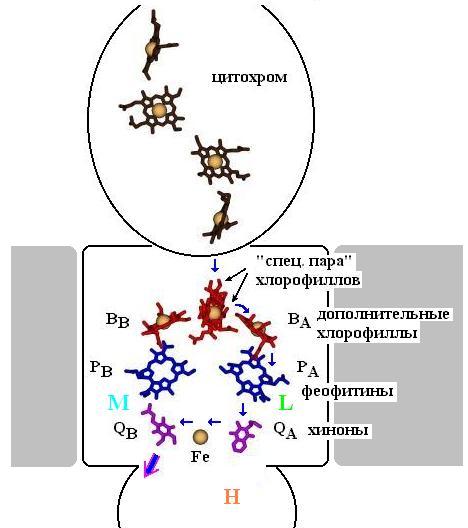
Рис.12-6. Схема расположения фотосинтетических пигментов в фотосинтетическом реакционном центре. Ось псевдосимметрии L и М субъединиц проходит через "специальную пару" хлорофиллов и ион Fe. Длинные "хвосты" пигментов "сбриты", чтобы не загромождать рисунок. Электрон преимущественно идет по правой (на рисунке) ветви пигментов, привязанной к субъединице L. Путь электрона отмечен синими стрелками. Большая фиолетово-синяя стрелка отмечает выход хинона, нагруженного двумя последовательно прибывшими электронами. Левая цепь не используется. Предполагается, что она использовалась в прошлом, а теперь осталась, как аппендикс.
Как же работает фотосинтетический реакционный центр?
Сначала квант света выбивает электрон из сидящей в белке "специальной пары" ("special pair") хлорофиллов (см. схему на Рис.12-6). Этот электрон мгновенно (менее чем за 10- 12 сек) перескакивает на соседний "добавочный" (accessory) хлорофилл, отстоящий на 3![]() от ближайшего хлорофилла "специальной пары", а затем — за ~10-10 сек, через феофитин РА, — на хинон (quinone) QA (именно QA, а не на QB!). Потом — уже за ~10-4 сек — электрон переходит на хинон QВ. Почему электрон идет столь кружным путем на хинон QВ, почему он не регистрируется (по электронным спектрам) на феофитине ВВ — еще неясно.
от ближайшего хлорофилла "специальной пары", а затем — за ~10-10 сек, через феофитин РА, — на хинон (quinone) QA (именно QA, а не на QB!). Потом — уже за ~10-4 сек — электрон переходит на хинон QВ. Почему электрон идет столь кружным путем на хинон QВ, почему он не регистрируется (по электронным спектрам) на феофитине ВВ — еще неясно.
Ушедший со "специальной пары" хлорофиллов электрон возмещается электроном, пришедшим с гема цитохрома. На этом завершается первый полуцикл реакции.
В результате второго аналогичного полуцикла, хинон QB нагружается вторым электроном и уходит с обоими электронами (дважды заряженный, он легче вырывается из мембраны), чтобы принять участие в дальнейшем фотосинтезе.
Итак, фотосинтетический реакционный центр переносит электроны из верхнего (на Рис.12-5, 12-6) в нижний компартмент — против создающейся, в результате, разности потенциалов между компартментами. Коэффициент полезного действия этого реакционного центра — около 50% (т. е. 50% поглощенного света превращается в энергию разделенных зарядов: вполне неплохо!).
Отметим два важных физических момента:
1) И хлорофиллы (Рис.12-7), и феофитины, и хиноны, и другие пигменты включают в себя системы "полуторных" (...-С=C-С=...), т. е. p-электронных связей. Иначе говоря, электронные облака в этих молекулах обобществлены (вследствие Полингова резонанса: ...С=С-... Û ...С-С=...), и электроны бегают по этим молекулам, как по кусочкам металла. Тем самым создается потенциальная яма для электронов, где они "делокализованы", т. е. могут смещаться на расстояния, значительно превышающие диаметр атома (кстати, именно делокализация электронов создает характерную окрашенность пигментов: локализованный в отдельной валентной связи электрон возбуждается коротковолновым ультрафиолетовым светом, а делокализованный электрон — "обычным", более длинноволновым светом).
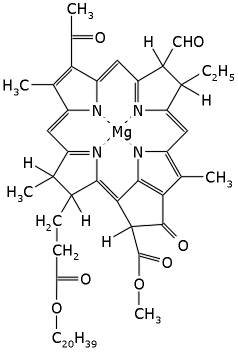
Рис.12-7. Молекула бактериохлорофилла.
2) Передача электрона с одного "кусочка металла" (пигмента) на другой не требует непосредственного контакта этих пигментов. Эта передача осуществляется путем квантового туннельного перехода (см. схему на Рис.12-8).

Рис.12-8. Схема, поясняющая туннельный эффект. Жирная линия показывает профиль потенциальной энергии U электрона. Пунктир — уровень полной (потенциальной + кинетической) энергии электрона. Тонкая линия (со штриховкой под ней) — плотность электронного облака r. Электрон первоначально сидит в левой яме (это его состояние и показано на рисунке), но край его облака (пусть и обладающего здесь очень малой плотностью) достигает правой ямы, так что электрон может со временем весь перетечь туда, если там энергия ниже. При глубине ямы в несколько электрон-вольт (характерной энергии, необходимой для ионизации молекулы) — характерное расстояние, на котором плотность его облака спадает на порядок, — около 1![]() .
.
Суть туннельного перехода (или, как говорят, "подбарьерного" перехода: ведь при этом электрон проходит как бы под энергетическим барьером), — в том, что, по законам квантовой механики, электрон (как и всякая частица, а в особенности — легкая частица) несколько "выступает" за пределы той потенциальной ямы, в которой он находится. "Потенциальной ямой" (областью низкой потенциальной энергии U) в данном случае служит та молекула (хлорофилл, феофитин и т. д.), на которой сидит электрон. Вне "ямы", в которой сидит этот электрон (Рис.12-8), его потенциальная энергия выше, чем его полная (потенциальная + кинетическая) энергия в яме. Если бы не квантовый эффект, этот дефицит энергии не давал бы электрону возможности высовываться из ямы. Квантовый же эффект приводит к тому, что волновая функция электрона (или, попросту, плотность электронного облака) "выплескивается" за потенциальную яму, — хотя величина этой плотности и очень быстро, экспоненциально спадает по мере удаления от "ямы".
Это — тот же квантовый эффект, что не позволяет электрону упасть на ядро: хоть это падение и уменьшило бы потенциальную энергию электрона, его кинетическая энергия возросла бы при этом еще больше. Дело здесь в том, что, при стремлении к нулю расстояния от электрона до ядра (Dx), — потенциальная энергия электростатического взаимодействия электрона с ядром стремится к минус бесконечности как 1/(Dx), а его кинетическая энергия, в силу принципа неопределенности Гайзенберга стремится (при Dx®0) к плюс бесконечности куда быстрее, — как 1/(Dx)2.
Действительно, согласно принципу Гайзенберга, неопределенность в скорости (Dv) и неопределенность в координате (Dx) частицы связаны соотношением mDvDx ~ ![]() , где
, где ![]() — постоянная Планка, а m — масса частицы. Иными словами, абсолютная величина скорости частицы (v) в яме шириной Dx составляет порядка
— постоянная Планка, а m — масса частицы. Иными словами, абсолютная величина скорости частицы (v) в яме шириной Dx составляет порядка ![]() /(mDx) — при полной неопределенности направления движения частицы в данный момент времени. Следовательно, кинетическая энергия частицы, E = mv2/2, есть величина порядка m[
/(mDx) — при полной неопределенности направления движения частицы в данный момент времени. Следовательно, кинетическая энергия частицы, E = mv2/2, есть величина порядка m[![]() /mDx)] 2 = (
/mDx)] 2 = (![]() 2/m)/Dx2.
2/m)/Dx2.
Так же обстоит дело и в случае электрона в потенциальной яме: если бы он ни на йоту не выступал бы за свою яму, его полная энергия была бы выше.
Поэтому электронное облако слегка "выплескивается" за потенциальную яму. При этом плотность его спадает экспоненциально — как и плотность электронного облака атома. Характерное расстояние, на котором плотность этого облака спадает на порядок (в 10 раз) — около 1![]() (это, как мы знаем, — характерный радиус атома).
(это, как мы знаем, — характерный радиус атома).
Значит, плотность электронного облака спадает в 1000 раз на расстоянии в 3 ![]() от "своей ямы". Это означает, что вероятность того, что за одно колебание электрон отойдет от "своей ямы" на 3
от "своей ямы". Это означает, что вероятность того, что за одно колебание электрон отойдет от "своей ямы" на 3 ![]() — порядка 10-3 (а на 5
— порядка 10-3 (а на 5 ![]() , — порядка 10-5, на 10
, — порядка 10-5, на 10 ![]() , — порядка 10-10 и т. д.). Электрон в "яме" (молекуле пигмента) совершает ~1015 колебаний в секунду (в видимом световом диапазоне частот: это хорошо видно по спектрам поглощения таких молекул). Значит, характерное время его перехода в другую "яму" (другую молекулу пигмента), отстоящую на 3
, — порядка 10-10 и т. д.). Электрон в "яме" (молекуле пигмента) совершает ~1015 колебаний в секунду (в видимом световом диапазоне частот: это хорошо видно по спектрам поглощения таких молекул). Значит, характерное время его перехода в другую "яму" (другую молекулу пигмента), отстоящую на 3 ![]() — порядка 10-12 сек; в яму, отстоящую на 5
— порядка 10-12 сек; в яму, отстоящую на 5 ![]() — порядка 10- 10 сек; в яму, отстоящую на 10
— порядка 10- 10 сек; в яму, отстоящую на 10 ![]() — 10-5 сек; а в яму, отстоящую на 15
— 10-5 сек; а в яму, отстоящую на 15 ![]() — порядка 1 сек. Такая зависимость скоростей переходов от расстояний согласуется с тем, что наблюдается в фотосинтетическом реакционном центре.
— порядка 1 сек. Такая зависимость скоростей переходов от расстояний согласуется с тем, что наблюдается в фотосинтетическом реакционном центре.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
